
Apparato uditivo umano
Apparato uditivo umano
ANATOMIA FUNZIONALE DELL’ORECCHIO
CARATTERISTICHE GENERALI
L’orecchio è suddiviso in tre porzioni: esterno, medio e interno, così dette per la loro disposizione topografica. Tutti e tre assolvono la funzione uditiva, ma solo l’orecchio interno contiene l’apparato vestibolare (che rileva la posizione della testa nello spazio).
Evoluzione: l’apparato vestibolare è la componente più antica dell’orecchio (deriva dai pesci). Con l’evoluzione e il passaggio dall’ambiente acquoso a quello terrestre, ha visto lo sviluppo dell’orecchio medio, affinché l’onda sonora non fosse smorzata.
L’orecchio, nelle sue porzioni, è in rapporto con l’osso temporale: il timpano è un’estroflessione dell’intestino primitivo, verso l’osso temporale in formazione.
Osso temporale: origina da quattro abbozzi fusi assieme, che danno vita alle varie porzioni.
- Squama ancestrale à squama e processo zigomatico (proc. Timpanoidale)
- Parte timpanica à meato acustico est. (canale acustico int.)
- Parte petromastoidea à proc. mastoideo, proc. Piramidale (rocca petrosa)
- Parte stiloidea à proc.stiloideo (origina dagli archi branchiali)
Prima di fondersi, i quattro abbozzi rimangono uniti da suture che nella vita del bambino ossificano diventando sinostosi. Rimangono comunque alcune fessure attraversate da nervi e vasi: petrotimpanica, petrosquamosa e timpanosquamosa.
L’orecchio è quasi tutto contenuto nella porzione timpanica e nella rocca petrosa (piramidale e mastoideo.
Sulla faccia anteriore del temporale vi è una piccola depressione per il ganglio semilunare sensitivo del trigemino. Sulla faccia inferiore sono invece il canale carotico e il foro giugulare.
LA PERCEZIONE UDITIVA
Trasduzione: processo che trasforma uno stimolo meccanico in un impulso elettrico tramite una cellula detta recettore. L’impulso si ha con la depolarizzazione. Il suono è percepito con questo meccanismo (cellule di corti nell’orecchio.
Il segnale per essere in grado di portare informazioni deve avere due caratteristiche: quantitativo (intensità dello stimolo) e qualitativo (tipo di stimolo).
Orecchio: distanziocettore che raccoglie, trasmette, modifica, analizza e amplifica le onde sonore e le rende impulso elettrico. E’ sensibile a differenze d’intensità, altezza e fase, percepisce velocemente la variazione di tali parametri. Rileva sorgente e distanza della fonte sonora.
ORECCHIO ESTERNO
COMPONENTI:
- Padiglione auricolare (o pinna)
- Meato acustico esterno (o canale uditivo esterno)
Padiglione: 6-7 cm x 3-4 cm. Accresce particolarmente nella vecchiaia e nell’infanzia.
Il rilievo esterno, la piega distale è l’elice, mentre l’antielice è più mediale e anteriore. Il trago (opposto alla porzione maggiore del timpano) e l’antitrago, chiudono in parte il meato. Tra elice e antielice c’è la fossa scafoidea. All’origine dell’antielice c’è la fossa triangolare. Al centro del padiglione c’è la conca, che si continua con il meato acustico esterno.
La faccia mediale dell’orecchio ha fosse e rilievi corrispondenti alla faccia laterale.
I rilievi servono ad eliminare i rumori di fondo: sfasando le onde sonore, creano treni d’onda per mettere in fase la voce umana.
L’angolo cefaloauricolare (con il cranio) si mantiene tra 5-25°.
Il padiglione è costituito da porzioni di cartilagine elastica che ne riprendono la forma.
Presenta legamenti intrinseci ed estrinseci, muscoli intrinseci (che determinano la forma dell’orecchio) ed estrinseci (per il suo movimento). I muscoli del padiglione sono parte dei muscoli mimici innervati dal n.faciale.
Nel lobulo vi è solo tessuto adiposo.
Irrorazione: - auricolare posteriore
- occipitale
- temporale posteriore (rami della carotide esterna).
Innervazione: - ramo agricolotemporale del trigemino
- n.vago (ramo auricolare) e n.glossofaringeo (conca dell’orecchio)
- rami del plesso cervicale.
Un’iperstimolazione vagale provoca: vomito, tosse, rallentamento del battito cardiaco, morte.
Un danno al padiglione provoca difficoltà nel discriminare chiaramente la voce umana e difficoltà nell’individuazione della sorgente sonora.
Meato acustico: Inizia in corrispondenza della conca e termina con la membrana del timpano (confine tra orecchio esterno e medio).
Lungo 2-3 cm. Il terzo lat. è cartilagineo ed è chiuso superiormente da connettivo fibroso. I due terzi mediali sono scavati nel temporale ed è chiuso superiormente dalla squama.
Porion: punto di repere. Punto più posterosuperiore del meato acustico.
Piano di Francoforte: piano passante per i due porion e per il margine inferiore delle due orbite.
E’ un canale che descrive una “S talica”(ruotata su due piani). Il percorso termina sulla membrana timpanica che, posta obliquamente, descrive col piano orizzontale un angolo di circa 55°. Nel bambino il canale è quasi rettilineo e la membrana è orizzontale. Lo specialista per osservare la membrana, tende l’elice in alto e all’esterno ( nel bambino in basso e indietro).
Istmo del canale: restringimento tra osso e cartilagine.
La cute riveste il padiglione, entra nel meato acustico fino a rivestire la parete laterale della membrana timpanica. La cute aderisce al periostio: vasi e nervi sono più comprimibili.
Ghiandole ceruminose: ghiandole sottocutanee lubrificanti del meato e della membrana, catturanti il pulviscolo atmosferico. Impediscono la lacerazione della cute.
Tragi: peli voluminosi per intrappolare gli insetti
Funzioni: - amplifica le onde sonore del parlato
- distribuisce la pressione sulla membrana timpanica
Innervazione e vascolarizzazione sono quasi le stesse del padiglione.
L’orecchio esterno contrae diversi rapporti:
- Fossa mandibolare e Articolazione TemporoMandibolare (ant.);
- Parotide (inferiormente);
- A.carotide esterna e N.faciale (dal foro stilomastoideo);
- Proc. Mastoideo e cellette mastoideee (posteriormente);
Patologie: Flogosi (otite esterna batterica, micotica o virale), neoplasie, atresia auris
ORECCHIO MEDIO
Componenti:
- membrana del timpano;
- cavo del timpano;
- martello, incudine, staffa
- cellette mastoidee (cavità piene d’aria, comunicanti tra loro e con i cavo del timpano);
- tuba uditiva ( di Eustachio).
Membrana : 1 cm x 0,8 cm. E’ inserita sull’orifizio mediale del meato acustico esterno. Ha forma alveolare. La superficie laterale è concava, quella mediale è convessa.
Nella porzione laterale, il punto di massima introflessione è detta ombelico o umbo. La stria malleolare è una striatura diretta in alto e in avanti a partire dall’umbo (corrisponde a ciò che si vede in trasparenza del manico del martello). La stria termina con due legamenti: i timpano malleolari ant. e post. per legare assieme la membrana all’orifizio del meato.
La membrana è costituita al 90% da pars tensa (la porzione tesa e rigida che vibra) e al 10% dalla pars flaccida (che non vibra).
Tracciando due rette, una passante per la stria malleolare e un’altra perpendicolare a questa ma passante per l’umbo, si individuano 4 quadranti: ant, post, sup, inf. Una membrana sana e correttamente tesa, presenta nel quadrante anteroinferiore un riflesso luminoso, il triangolo luminoso del Politzer (indicatore di corretta tensione).
il quadrante posteroinferiore è quello scelto per le perforazioni in sede operatoria.
La membrana timpanica è costituita da tre strati:
1) cutaneo (sulla faccia laterale)
2) connettivale (intermedio)
3) mucoso (sulla faccia del cavo timpanico).
Secondo una ulteriore suddivisione, la porzione centrale è la vera membrana timpanica: il connettivo centrale è elastico, con andamento orientato su due strati (laterale e mediale). Lo strato laterale ha tre tipi di fibre con tre andamenti:
- Fibre radiate: dal manico del martello si inseriscono sui margini della membrana, disponendosi a raggiera.
- Fibre paraboliche: dal manico del martello, con archi più o meno arcuati, vanno sulle fibre del martello
- Fibre semilunari: originano e terminano sul margine della membrana.
Lo strato mediale ha un unico tipo di fibre, quelle circolari, che hanno origine ed inserzione sul manico del martello. Il manico del martello è parte della membrana timpanica: la sua vibrazione, mette in vibrazione il martello. Il manico può aumentare o diminuire concavità o convessità per rispondere meglio ai suoni: se è molto teso e convesso riceve in maniera ottimale suoni acuti e viceversa.
Tutte queste fibre sono presenti solo nella pars densa. Nella pars flaccida ci sono solo poche fibre elastiche.
timpano
- ANT à asportata nell’immagine
- LAT à occupata dalla membrana del timpano
- MED à separa il cavo del timpano dall’orecchio interno
- INF à pavimento del cavo
- SUP à tetto, vede sulla sua superficie anche la corda del timpano
- POST à è in profondità, ospita l’aditus ad antrum, un orifizio che immette nell’antro mastoideo.
Parete laterale e mediale sono collegate dalla catena dei tre ossicini (martello, incudine e staffa).
In realtà la cavità è molto irregolare: prendendo come riferimenti i limiti, superiore e inferiore del meato acustico esterno si descrivono due ulteriori cavità. Sopra al meato acustico si trova il recesso epitimpanico (epitimpano o soggetta degli ossicini) e contiene martello e incudine; la porzione compresa tra i margini del meato acustico è il cavo del timpano vero e proprio; la porzione al di sotto del meato acustico è detto recesso ipotimpanico (o ipotimpano). L’epitimpano ha un recesso sulla sua faccia posteriore: l’antro mastoideo.
Martello: ha una testa e più processi. Il manico è un processo che si dirige in basso obliquamente. Il processo breve è corto e tozzo. Il processo anteriore è lungo e affusolato. Sulla faccia mediale della testa, c’è la faccetta articolare per l’incudine.
Incudine: ha un corpo centrale con la faccetta articolare per il martello laterale. Ha un processo breve (corto e tozzo) e un processo lungo (affusolato) al cui termine si trova il processo lenticolare con la faccetta per l’articolazione con la staffa. I processi fissano, tramite legamenti, gli ossicini alle pareti del cavo.
Staffa: ha una base che stacca un arco anteriore e un arco posteriore, che convergono nella testa della staffa, che presenta una faccetta articolare per l’incudine. La base è articolata con la finestra ovale, un orifizio sulla parete mediale del cavo (non c’è membrana). Il movimento della staffa, non è la compressione, ma la rotazione attorno all’asse maggiore della base.
Le articolazioni tra queste ossa sono diartrosi, mobilli, con cartilagini e capsule articolari. L’articolazione tra martello e incudine presto ossifica in maniera funzionale alla trasmissione della vibrazione. La mucosa riveste il cavo del timpano e gli ossicini con epitelio cubico.
Tetto: è la porzione ossea che copre il cavo del timpano. È obliquo e orientato inferoanteriormente. E’ detto tegmen timpani: separa il cavo del timpano dalla fossa cranica media.
Pavimento: è più spesso rispetto al tetto. Trovandoci nella piramide del temporale, separa il cavo del timpano dal bulbo superiore della vena giugulare interna.
Parete Anteriore: si apre la tuba uditiva di Eustachio.
Tuba uditiva: qui ha il suo orifizio timpanico e sbocca con l’orifizio faringeo, sulla parete laterale della rinofaringe. E’ un canale di 3 cm, osseo prossimalmente e cartilagineo distalmente, obliquo lateromedialmente. La architettura cartilaginea è su 3 lati, quella laterale è di connettivo fibroso: il canale così, non è sempre beante. Sul torus tubarius sono inseriti: il m.elevatore e tensore del velo palatino, m.salpingofaringeo.
Parete Posteriore: accoglie un canale osseo al cui interno decorre il n.faciale. Questo nervo nel suo percorso stacca la corda del timpano (componente parasimpatica gangliare per le ghiandole e componente sensitiva speciale per la papille fungiformi del corpo della lingua).
Qui è anche accolto il muscolo stapedio, che col suo tendine si aggancia alla testa della staffa (è innervato dal n.faciale). Risponde involontariamente ai riflessi: quando lo stapedio è contratto, la staffa non è più in grado di comprimere l’orecchio interno; questo è un meccanismo di difesa.
Corda del timpano: attraversa tutto il cavo e fuoriesce dalla fessura petrotimpanica (sulla faccia inferiore del temporale).
Parete Mediale: ha il promontorio, un rilievo nel cavo del timpano che corrisponde al giro basale della chiocciola ossea nell’orecchio interno. Il plesso nervoso e sanguigno timpanici si trovano su tale promontorio. Ci sono due orifizi principali:
- finestra ovale, occupata dalla base della staffa, conduce all’orecchio interno;
- finestra rotonda, anteroinferiore, chiusa dal connettivo elastico della membrana secondaria del timpano (nel vivente). Tale finestra è uno sfogo per la pressione esercitata dalla staffa nell’orecchio interno (fenomeno dovuto all’incomprimibilità dei liquidi).
Superiormente alla tuba uditiva, c’è il canale che accoglie il muscolo tensore del timpano (striato, innervato da un ramo del n.trigemino). Dall’orifizio di tale canale ne fuoriesce solamente il tendine, che inserendosi sul collo del martello, può trascinarlo medialmente regolando la convessità della membrana timpanica. Risponde involontariamente ai riflessi: appena arriva il primo impulso sonoro, garantisce una risposta ottimale della membrana
Inferiormente a tale parete scorre l’arteria carotide interna nel canale carotico.
Il canale del n. faciale connette orecchio interno e medio e descrive un “ginocchio” in prossimità del ganglio sensitivo. Il n.faciale (con le sue componenti speciale e motoria) fuoriesce dal foro stilomastoideo.
Iperacusia: i suoni sono avvertiti molto più forti a causa del danneggiamento dello stapedio che regola la staffa.
Blocco della catena ossiculare: per evitare danni all’orecchio interno, stapedio e tensore del timpano possono agire assieme arrestando gli ossicini.
Il cavo del timpano ha la funzione di trasmettere non smorzato, anzi amplificato il suono. Poiché la finestra ovale è ampia dieci volte in meno della membrana del timpano, e su di essa viene esercitata la stessa forza che si esercitava nel meato acustico esterno, la pressione è ovviamente maggiore. La catena degli ossicini poi, è una leva vantaggiosa: aumenta la forza con cui lavora la staffa.
Cellette Mastoidee: cavità comunicanti piene d’aria, all’interno del processo mastoideo. E’ in comunicazione con l’antro timpanico (i cm3 ). La mastoidite è l’infiammazione della mucosa delle cellette mastoidee (poco curabile perché è una zona poco vascolarizzata in cui gli antibiotici non hanno effetto).
Superiormente al meato acustico esterno, sull’osso temporale vi è l’area cribrosa di Mac Ewan, da cui si può accedere all’orecchio interno, tramite l’antro mastoideo.
ORECCHIO INTERNO
Si trova nella piramide del temporale. Contiene sia la funzione acustica che quella vestibolare. E’ chiusa nel vivente e in comunicazione col cavo del timpano tramite le finestre ovali e rotonda.
Si parla di labirinto osseo, una serie di cavità in comunicazione (spesso ci è mostrato il calco in gesso), al cui interno risiede il labirinto membranoso. Il labirinto osseo è costituito da:
- vestibolo (porzione centrale allargata);
- canali semicircolari (canali ossei che a partire dalla porzione posteriore dal vestibolo, descrivono degli archi di cerchio che tornano in esso);
- coclea o chiocciola ossea (porzione convoluta anteriore al vestibolo);
- acquedotto del vestibolo (canalicolo laterale)
- acquedotto della coclea (canalicolo che parte inferiormente alla base della coclea).
Vestibolo e canali semicircolari contengono l’organo dell’equilibrio, la coclea quello dell’udito.
Semicircolari
Coclea: cavità avvolta su se stessa. La porzione centrale attorno cui si scava il canale è detta modiolo (tronco di cono con apice sull’apice della coclea e base sul fondo del meato acustico interno). Attorno al modaiolo si descrivono due giri e mezzo di canale. Dal modiolo si stacca la lamina spirale ossea, laminetta che divide il canale (per chiuderlo completamente sarà coadiuvata da una lamina membranosa) in due compartimenti che procedono adiacenti: la scala vestibolare (superiore e in rapporto con la finestra ovale) e la scala timpanica (inferiore e in rapporto con la finestra rotonda).
All’interno del labirinto osseo c’è il labirinto membranoso, una serie di vescichette comunicanti variamente contenute. Ha diverse porzioni:
- Utricolo (vescichetta maggiore e posteriore) e sacculo (vescichetta minore e anteriore) collegate dal dotto reuniente. Sono contenute nel vestibolo osseo;
- Canali semicircolari membranosi (contenuti in quelli ossei);
- Condotto cocleare membranoso (contiene perilinfa, coadiuva la lamina spirale ossea ed è contenuto nella coclea ossea);
- Dotto endolinfatico (a partire dal dotto reuniente, termina con il sacco endolinfatico o allargato. E’ accolto nell’acquedotto del vestibolo).
Il labirinto membranoso occupa solo un sesto del labirinto osseo, a cui si ancora tramite tralci connettivali. Il resto è occupato dal liquido detto perilinfa. All’interno del labirinto membranoso scorre invece endolinfa.
L’architettura esterna è di connettivo fibroso (che consente di fissarsi all’osso e non fluttuare). La cavità interna è rivestita da epitelio pavimentoso monostratificato (epitelio sensoriale in alcuni punti per trasformare l’impulso meccanico in elettrico).
Endolinfa: extracellulare, con composizione ionica simile a quella cellulare (ricca di K+ e povera di Ca++ e Na+). E’ prodotta da:
- stria vascolare;
- membrana del Reissner;
- dark cells (nella porzione vestibolare, mantengono le differenze di concentrazione).
E’ drenata dal dotto e dal sacco endolinfatici (e poi nei seni venosi della dura madre). Deve rimanere costante in quantità.
Perilinfa: composizione simile ai liquidi extracellulari (tanti na+ e meno K+). Prodotta come trasudato dei vasi sanguigni di vestibolo e coclea. E’ drenato dall’acquedotto cocleare che immette nei seni venosi della dura madre.
EPITELIO SENSORIALE
L’epitelio sensoriale del labirinto membranoso ha due tipi di cellule:
- Cellule ciliate sensoriali (uditive o vestibolari);
- Cellule di sostegno (simili alle cellule gliali, veicolano gli impulsi isolando elettricamente due cellule sensoriali adiacenti).
In ogni orecchio ci sono 20.000 cellule acustiche e 160000 cellule vestibolari.
Le cellule ciliate sensoriali sono dotate, al loro apice, di estroflessioni: il chinociglio (un vero ciglio vibratile) e più stereociglia (estroflessioni del plasmalemma non in grado di muoversi).
Le stereociglia sono disposte in ordine decrescente a partire dal chinociglio e piegandosi sono in grado di generare corrente elettrica. Sono collegate tra loro e al chinociglio da filamenti proteici del citoscheletro, i tip-link (un filamento di caderina 23): quando si muove attivamente il chinociglio, le stereociglia si muovono passivamente. Si parla di pelo acustico.
Il tip-link collega due canali ionici, di due stereociglia diverse, per il passaggio di ioni K+. Trasferisce la tensione di flessione: come una molla al momento della tensione trasferisce l’energia alle estremità aprendo i canali. Se la flessione ha la direzione di quella del chinociglio allora i canali si aprono; nel caso inverso si chiudono. Se il tip-link è distrutto o manca la cellula ciliata non funziona.
Quando un canale viene aperto, i K+ dell’endolinfa penetrano nella cellula e il potenziale di membrana (fisiologicamente negativo) viene alterato. La cellula viene così depolarizzata (le cellule si depolarizzano a differenze di potenziale diverse, -40v o –80v): si aprono i canali del Ca++ a controllo di voltaggio. Il Ca++ diffonde nella cellula e spinge cambiamenti conformazionali del citoscheletro che fanno fondere le vescicole piene di neurotrasmettitori (glutammato) nello spazio intersinaptico innescando il segnale elettrico. Il Ca++ nella cellula attiva la pompa Na+/K+ ATPasi che riporta la situazione ionica alla normalità.
Nonostante questo meccanismo si attivi solo con il movimento delle stereociglia, in condizioni di riposo, alcuni canali per il K+ possono rimanere aperti che invieranno un certo numero di segnali (al massimo varierà la velocità di invio di tali segnali), che non è detto destino la soglia di attenzione.
In assenza del chinociglio le stereociglia si muovono lo stesso.
Le cellule sensoriali possono variare per:
- lunghezza e geometria delle stereociglia;
- diversa flessibilità, densità e distanza delle stereociglia;
- numero diverso di canali per il K+;
- diversa struttura del tip-link;
- diversità dei canali per il K+.
Alla base delle cellule sensoriali arrivano terminazioni nervose che hanno due modi di sinaptare:
- terminazioni afferenti (dendrite verso i centri superiori);
- Terminazioni efferenti (assone dai centri superiori).
APPARATO VESTIBOLARE
Gli epiteli sensoriali dell’apparato vestibolare sono dette macule e si trovano nel sacculo e nell’utricolo. La macula del sacculo è verticale, quella dell’utricolo è orizzontale, sono disposte tra loro perpendicolarmente.
L’epitelio sensoriale nei canali semicircolari membranosi è sito nella regione dell’ampolla e costituisce le creste ampollari (tra loro perpendicolari).
Ogni orecchio interno contiene dunque 5 porzioni recettoriali.
Ampollari
- Tipo 1 (a fiasco): contraggono sinapsi afferenti (asso dendritica) caliciformi, sono veloci ed efficienti. La terminazione sinaptica efferente sinapta con quella afferente.
- Tipo 2 (a bastoncello): contraggono sinapsi afferenti uniformi normali.
In entrambi i casi i centri superiori regolano la terminazione afferente.
Queste cellule sono attivate ogni volta che con la testa eseguiamo accelerazioni angolari
(rotazioni) attorno ad un asse. La rotazione in una data direzione fa si che la cupola gelatinosa e l’endolinfa rimangano indietro, in modo da piegare i peli acustici in direzione opposta al
movimento della testa. L’effetto sulle stereociglia di ogni canale semicircolare e di ogni orecchio sarà diverso: le 6 creste ampollari invieranno segnali diversi.
I centri superiori regolano l’equilibrio dinamico, integrano tali segnali e coordinano il movimento degli occhi con la rotazione della testa (riflessi vestibolo-oculari).
Macula: porzioni sensoriali simili alle creste ampollari, con cellule di sostegno pseudostratificate e peli acustici. Questi sono inglobati nella membrana otolitica che contiene gli otoconi (sali di calcio nella matrice mucopolisaccaridica) ed è più pesante e densa dell’endolinfa circostante. Le cellule sono attivate da accelerazioni lineari o dalla forza di gravità: piegando la testa in basso, la membrana trascina i peli acustici in basso più velocemente, provocando la piega.
In ogni macula, si possono distinguere due gruppi di cellule che si comportano in modo indipendente: ogni impulso meccanico attiva dunque non 4 ma 8 impulsi elettrici. I centri superiori dalle macule ricavano informazioni per l’equilibrio statico e per la postura.
Ogni movimento dunque attiva 14 gruppi di cellule e quindi 14 impulsi, che verranno integrati dai centri superiori. I dendriti che innervano le cellule ampollari hanno il soma nel ganglio vestibolare o dello Scarpa, sul fondo del meato acustico interno (neuroni unipolari, non pseudo). Il tutto è mielinizzato e il n.vestibolare sarà formato dalla confluenza degli assoni, con direzione nuclei vestibolari del ponte e del cervelletto.
Dai nuclei vestibolari, le fibre si dirigeranno alla corteccia telencefalica, tramite due vie:
- Vie vestibolari centrali: dirette al telencefalo (nell’area 5 del lobo parietale), questo serve ad avere coscienza dell’orientamento della testa nello spazio per regolare i muscoli antigravitari in modo da non cadere.
- Vie vestibolospinali: nel midollo spinale.
APPARATO UDITIVO
E’ contenuto unicamente nella chiocciola ossea. Grazie al dotto cocleare membranoso, la coclea ossea è suddivisa completamente nelle due scale. Il dotto cocleare fa si che si possano individuare in ogni sezione tre rampe che ruotano attorno al modaiolo:
- dotto vestibolare: contenente perilinfa;
- dotto timpanico: contenente perilinfa;
- dotto cocleare: tra i due precedenti, contenente endolinfa (detta anche scala media).
La scala media è più corta del canale osseo: all’apice della chiocciola ossea, le altre due scale sono in continuità tramite l’elicotremo (di dimensioni variabili).
Il volume della scala media, una volta adesa alla parete ossea, varia e assume sezione triangolare. Si modificano la superficie connettivale e l’epitelio di rivestimento: alla base del triangolo si ha infatti epitelio sensoriale. Tutte queste strutture sono attraversate da vasi e nervi.
Lato esterno: Sul lato esterno della coclea membranosa, il connettivo fibroso si è fuso con il periostio e l’epitelio di rivestimento diventa cilindrico pseudostratificato detto stria vascolare (specializzato nel produrre endolifa a partire dai vasi che scorrono nelle vicinanze).
Lato obliquo: rimane libero e l’epitelio rimane pavimentoso monostratificato. Questo lato è anche detto membrana vestibolare o del Reissner. Separa la scala vestibolare dalla scala media e le loro linfe. Partecipa a mantenere la differenza di concentrazioni di endolinfa e perilinfa (tramite delle pompe sulla sua superficie). Collabora a produrre endolinfa.
Il lato inferiore è la porzione che subisce i maggiori cambiamenti, perciò lo dividiamo in due zone:
Mediale: prende inserzione con la lamina spirale. Nel tratto della base è fissata all’osso: il connettivo si fonde col periostio e si ispessisce nel lembo spirale. Il margine superiore di tale lembo si introflette nei denti acustici che contengono le cellule dei denti acustici (cubiche modificate). Tali cellule mantengono la produzione della membrana tectoria. Tale membrana (ovoidale e allungata, in sezione verticale) ha matrice simile alla cupola gelatinosa, con qualche traccia di fibra collagene. Sporge lateralmente toccando l’organo recettoriale del Corti.
Laterale: tale porzione è tesa tra la lamina spirale ossea e la parete ossea della coclea. La componente connettivale è andata a formare la membrana basilare, mentre la componente epiteliale è andata a costituire l’organo recettoriale del Corti.
Membrana basilare: modificazione del tessuto connettivo. Si allarga e si assottiglia passando dal giro basale a quello apicale, arricchendosi di fibre elastiche (all’apice è molto flessibile). In base al tipo di stimolo (acuto o grave), viene messa in vibrazione, selettivamente, una porzione diversa di tale membrana.
Organo recettoriale del Corti: cellule epiteliali modificate. Si appoggia e origina dalla membrana basilare. Contiene sia cellule di sostegno (con funzione trofica o legate alle cellule acustiche) che sensoriali (non troppo diverse dalle cellule dell’apparato vestibolare, funzionano come visto in precedenza).
Cellule di sostegno: di tipo diverso a partire dal margine mediale. Medialmente ci sono i pilastri interni e i pilastri esterni, con una base larga, un lungo collo e una testa dilatata. Sono disposti ognuno in una singola riga. Le basi divergono ma la testa del pilastro interno accoglie quella del pilastro esterno descrivendo il tunnel del Corti. Fungono da isolanti.
Deiter’s cells: disposte in 3-5 righe. Hanno forma di clava, con un collo molto sottile e una testa schiacciata (cuticolo quadrangolare).
Henser’s cells: disposte in 3-5 righr. Hanno forma cilindrica, danno consistenza all’organo e si fondono con la membrana tectoria.
Claudius’s cells: cubiche, le più laterali, di sostegno e rivestimento.
Cellule acustiche interne: si trovano tra i colli dei pilastri interni. Hanno il chinociglio e le stereociglia. Sono orientate a formare una “U”. Sono circa 3500 con 120 stereociglia. Sono tozze
Cellule acustiche esterne: sparse ordinatamente in 3-5 file tra le deiter’s cells. Non hanno il chinociglio. Possono far contrarre le stereociglia modificandone l’altezza in seguito a impulsi dai centri superiori. Sono orientate a formare una “V”. Sono 20000 con 50 stereociglia. Sono più regolari ed affusolate
Le sinapsi delle due cellule non sono molto diverse, in quanto ricevono entrambe sinapsi afferenti (verso i centri superiori) e sinapsi efferenti (dai centri superiori).
Cellule con stereociglia più corte e rigide sono nei giri basali (freq. Acute), le stereociglia lunghe e affusolate sono nei giri apicali (freq. Basse).
Le stereociglia delle cellule acustiche non sono completamente inglobate nella membrana tectoria, ci arriva solo la porzione apicale di quelle più lunghe. Le cellule acustiche interne non la toccano mai.
la base delle cellule acustiche è raggiunta dal nervo cocleare con fibre miste: le afferenti originano da un ganglio, quelle efferenti dai centri superiori.
Trasduzione: Il meccanismo di trasduzione del segnale acustico è complesso.
La compressione dell’aria da parte dell’onda si trasmette dalla membrana, alla catena degli ossicini, alla perilinfa dell’orecchio interno. La finestra ovale comunica con la scala vestibolare: da qui passa l’onda, fino a giungere nell’elicotremo, nella scala timpanica, fino a trasmettere il suo impulso meccanico alla membrana della finestra rotonda che si estroflette verso il cavo del timpano.
Nella scala timpanica, l’onda metterà in vibrazione la membrana basilare del dotto coclearie membranoso, in uno dei livelli di flessibilità compatibile con le caratteristiche dell’onda. La membrana in vibrazione solleverà tutto l’organo del Corti e le stereociglia saranno più o meno inglobate nella membrana tectoria e verranno più o meno piegate.
Le cellule acustiche esterne invieranno un segnale ai centri superiori per modificare e regolare la forma delle cellule esterne stesse. La vibrazione dell’intero organo e i movimenti in altezza delle cellule, mettono in moto l’endolinfa. L’endolinfa piegherà le cellule acustiche interne attivandole: queste invieranno il vero segnale.
Cortilinfa: liquido ottenuto dalla miscela di endolinfa e perilinfa, dedita all’attivazione ottimale delle cellule acustiche.
Tonotopicità: fenomeno che caratterizza le vie acustiche e grazie al quale ogni punto della chiocciola membranosa risponde in maniera migliore ad una data frequenza. Generalmente i suoni acuti sono percepiti meglio dalla zona basale, quelli gravi dalla zona apicale. Vi sarà poi un gruppo di neuroni efferenti che trasporteranno l’impulso di quella determinata frequenza. I centri superiori terranno conto dell’impulso ricevuto unicamente da quel punto.
Innervazione: alla base delle cellule acustiche arrivano i dendriti del nervo coclearie.Le fibre emergono dall’estremità laterale dei foramina nervina (piccoli fori posti medialmente all’organo del corti, sulla membrana basilare) e raggiungono le cellule acustiche.
Dalle cellule acustiche, le fibre entrano nei foramina nervina, penetrano nella lamina spirale ossea e poi nel modiolo e raggiungono il ganglio spirale o del Corti (così detto perché è allungato e segue i giri della coclea). Tale ganglio è accolto dove la lamina si stacca dal modaiolo, nel canale di Rosental. Dal ganglio del Corti originano fibre afferenti (prolungamenti massonici)che attraversano i canalini del modiolo fino alla base. Filamenti del nervo cocleare fuoriescono dai forami dell’area cribrosa sul fondo del meato acustico interno.
Il ganglio spirale contiene somi di neuroni sensitivi di due tipi:
- TIPO I: (90-95%), bipolari, con guaina mielinica, che innervano le cellule acustiche interne con un rapporto di 10:1, cioè di 10 neuroni per una cellula acustica. Innervazione convergente;
- TIPO II: (5-10%), pseudounipolari, più piccoli. Sono diretti alle cellule acustiche esterne. Ogni neurone innerva fino a 10 cellule acustiche. Innervazione divergente.
Le fibre per le cellule acustiche interne possono regolare il segnale in modo da selezionare i suoni su cui poniamo attenzione (possiamo privilegiare certi tipi di suono).
Il n. acustico termina nei nuclei cocleari che rimandano alla corteccia uditiva. La via acustica decussa in vario modo: la corteccia di ogni lato riceve le informazioni da entrambe le orecchie. Un danno alle vie acustiche da difficoltà discriminativi della sorgente sonora.
SORDITA’
L’orecchio ha due tipi di funzionamento: meccanico ed elettrico. L’eziologia della sordità è dunque di due tipi: meccanica o nervosa.
Il danno nervoso può essere legato a cellule nervose sensoriali danneggiate da farmaci, soprattutto quelli ototossici (c.esterne più sensibili delle interne).
Il danno meccanico può essere legato a un danno della membrana timpanica o della catena ossiculare. Una membrana timpanica danneggiata in parte, a seconda dei casi può rigenerarsi. In caso contrario si opera una miringoplastica, una ricostruzione fatta usando una porzione di fascia del temporale. La riparazione della catena degli ossicini è fatta tramite due possibili operazioni: stapedectomia e stapedetomia.
Un ulteriore danneggiamento può provenire da traumi acustici: l’esposizione prolungata a suoni di elevata intensità (>90dB) danneggia sia le cellule sensoriali che i neuroni afferenti. La perdita dell’udito può essere temporanea o permanente. Le cellule acustiche (essendo nervose) possono rigenerarsi nel caso in cui non sia stato danneggiato il soma).
Il glutammato è neurotossico perché aumenta la concentrazione di Ca++, nocivo per le cellule,ad alte concentrazioni: il danno alle sinapsi afferenti si sviluppa in una fase acuta ed una tardiva.
VASCOLARIZZAZIONE E DRENAGGIO LINFATICO
Vascolarizzazione arteriosa
Arterie vertebrali à Arteria basilare à Arteria cerebellare anteriore inferiore à Arterie del labirinto (Vestibolare anteriore, Coclearie, Vestibolo cocleare).
Vascolarizzazione venosa
Seno petroso superiore à Seno traverso à Vena giugulare interna
Drenaggio linfatico
Orecchio esterno:• linfonodi parotidei, mastoidei, cervicali profondi
Orecchio medio: • linfonodi parotidei, retrofaringei, cervicali profondi
Orecchio interno: • non possiede vasi linfatici
Fonte: http://thenextdoctor.files.wordpress.com/2010/12/anatomia-funzionale-dell.doc
Apparato uditivo umano
ACUSTICA PSICOFISICA
Presentazione
Il fine di questa lezione è capire il funzionamento del sistema uditivo umano, in modo da prevedere la sensazione uditiva in funzione dello stimolo che la provoca. Conoscere l’udito umano è utile per la progettazione e la costruzione di sistemi acustici dal funzionamento dell’impianto stereo domestico, all’acustica di una grande sala.
La sensazione sonora
La risposta dell’orecchio umano ad uno stimolo sonoro è in generale un fenomeno molto soggettivo: essa infatti dipende da diversi fattori, come il deterioramento del sistema uditivo stesso dovuto a prolungate sollecitazioni. Un segnale sonoro è caratterizzato da due grandezze fondamentali: la pressione, legata all’intensità dell’onda che trasporta il segnale, e la frequenza. Sottoponendo individui otologicamente normali a diversi stimoli sonori, dei quali vengono variate pressione e frequenza, e analizzando le risposte uditive, è possibile tracciare un grafico, detto diagramma di sensazione, che mostri quali suoni sono percepibili dall’uomo.

Figura 1: Diagramma di sensazione
Il limite inferiore del grafico, la soglia di udibilità, rappresenta le pressioni minime, alle diverse frequenze, che vengono percepite dall’uomo; il limite superiore, la soglia del dolore, indica la massima intensità sonora che non provoca dolore; fra queste due linee si estende l’area della sensazione uditiva che contiene tutti i suoni udibili.
I ricercatori Fletcher e Munson, utilizzando un diverso approccio al problema, elaborarono negli anni ’30 un diagramma più completo, basato sulle curve isofoniche. Tali curve rappresentano il livello di pressione che deve avere un suono, alle diverse frequenze, per provocare la stessa sensazione. Il procedimento che adottarono era il seguente: un ascoltatore è sottoposto ad un suono puro, generato da un’onda piana sinusoidale con pressione e velocità in fase, e in seguito ad un suono di riferimento con frequenza 1000Hz; regolando l’intensità del suono di riferimento in modo che le due sensazioni corrispondano, si stabilisce a quale curva appartiene la coppia di valori pressione-frequenza del primo suono.
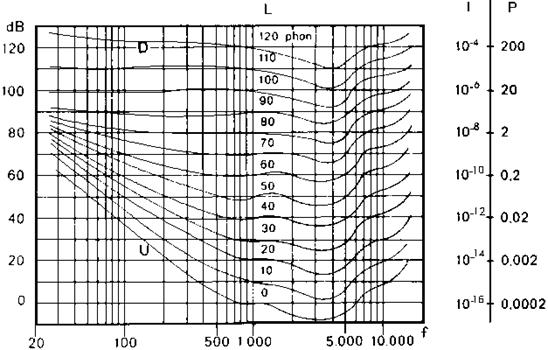
Figura 2: Audiogramma normale
Il nome “audiogramma normale” dato da Fletcher e Munson al grafico ottenuto non è propriamente corretto, perché le condizioni sotto le quali è stato ottenuto (purezza del suono, onde sinusoidali piane) non sono quasi mai confrontabili con la realtà. L’importanza di tale grafico è comunque considerevole, in quanto è utilizzato per valutare le misure effettuate con sistemi che hanno una risposta uguale a tutte le frequenze; su di esso si basano inoltre le normative che stabiliscono per legge il livello sonoro limite al quale può essere sottoposto l’uomo.
La scala dei Decibel
Per comprendere meglio tale grafico è utile definire una scala per i livelli sonori. Lo scienziato statunitense Graham Bell (1847-1922) osservò che la sensazione sonora, alla frequenza campione di 1000 Hz, raddoppia se l’intensità del suono che la provoca cresce di un fattore pari a circa 3.16 @ 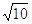 . Tale valore è naturalmente approssimato, in quanto la risposta ad una variazione in pressione dipende dalle caratteristiche dell’onda sonora, ma risulta essere abbastanza preciso. In particolare tale valore convinse Bell ad usare una scala logaritmica per misurare la sensazione sonora: scegliendo ad esempio una scala arbitraria alle varie pressioni si avrebbero i seguenti risultati
. Tale valore è naturalmente approssimato, in quanto la risposta ad una variazione in pressione dipende dalle caratteristiche dell’onda sonora, ma risulta essere abbastanza preciso. In particolare tale valore convinse Bell ad usare una scala logaritmica per misurare la sensazione sonora: scegliendo ad esempio una scala arbitraria alle varie pressioni si avrebbero i seguenti risultati
Pressione sonora (Pa) |
Sensazione (S) |
0.01 |
1 |
0.0316 |
2 |
0.1 |
3 |
0.316 |
4 |
1 |
5 |
dove l’aumento di S di un’unità indica il raddoppio della sensazione.
Bell definì quindi la sensazione sonora come:

utilizzando come unità di misura per tale grandezza il Bel [B] (dove lg indica il logaritmo in base dieci). Si può notare come le pressioni usate non siano quelle massime associate all’onda, ma il valor medio efficace (RMS) mediato su un periodo, in quanto tale valore è più semplice da calcolare. La quantità P0 è la pressione di riferimento, fissata al valore  Pa, che corrisponde al più basso suono udibile avente frequenza 1000 Hz. I valori ottenuti con tale formula corrispondono bene a quelli trovati sperimentalmente, ma tale scala si rivelò presto troppo grossolana: è per questo che ancora oggi si usano i suoi sottomultipli, in particolare il Decibel [dB]. Il Decibel non è una vera e propria unità di misura, ma indica il livello della grandezza al quale è riferito:
Pa, che corrisponde al più basso suono udibile avente frequenza 1000 Hz. I valori ottenuti con tale formula corrispondono bene a quelli trovati sperimentalmente, ma tale scala si rivelò presto troppo grossolana: è per questo che ancora oggi si usano i suoi sottomultipli, in particolare il Decibel [dB]. Il Decibel non è una vera e propria unità di misura, ma indica il livello della grandezza al quale è riferito:

il risultato di tale espressione è quindi il livello di pressione associato al suono.
Il Decibel viene riferito a qualsiasi grandezza di cui sia necessario avere una scala logaritmica; ad esempio le scale usate per indicare il volume di molti stereo sono espresse in decibel negativi: esse misurano il livello di attenuazione del segnale sonoro originario. Un’altra caratteristica importante del decibel risiede nella sua semplicità pratica ai fini del calcolo. Se, ad esempio, due suoni hanno una differenza nel livello della pressione pari a 6dB, attraverso semplici calcoli si può risalire alla differenza di pressione sonora che li distingue:

Se il segnale sonoro è trasmesso da un’onda piana sinusoidale con velocità e pressione in fase, è utile definire un livello per tutte le sue grandezze caratteristiche. Si definisce quindi il Livello di velocità:
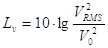
che indica la velocità dell’onda sonora rispetto alla velocità di riferimento; quest’ultimo valore si ricava facilmente ricordando che  :
:

Il Livello di intensità sonora:
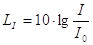
dove il valore di riferimento è dato da:
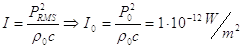
Infine il Livello di densità sonora:
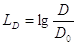
dove il valore di riferimento è dato da:
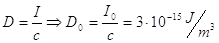
Utilizzando i livelli così definiti per analizzare un’onda piana sinusoidale si ottiene:
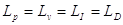
Somma di segnali sonori
Fino ad ora abbiamo analizzato segnali sonori puri caratterizzati da onde piane sinusoidali: nella realtà nessun segnale sonoro si presenta sotto tale forma. Vedremo più avanti che ogni onda, per quanto complicata essa sia, può essere scomposta nella somma di tante onde sinusoidali ognuna con una caratteristica intensità e fase. E’ quindi importante vedere il comportamento dei vari livelli sonori quando vengono sommati due o più toni puri.
La somma di due segnali si dice coerente quando questi sono identici e in fase: in tal caso il segnale risultante ha per pressione la somma delle pressioni delle sorgenti. Il livello totale risulta quindi essere:

la somma di due segnali a 50 dB risulta quindi di 56 dB.
La somma di due segnali si dice incoerente in tutti gli altri casi: non è detto che le pressioni si sommino aritmeticamente, poiché vi possono essere momenti in cui due picchi dell’onda si sommano o altri in cui un picco e una valle si annullano. In questi casi si ricorre al principio di conservazione dell’energia: l’intensità sonora dell’onda risultante è data dalla somma delle intensità delle sorgenti. Il livello totale risulta quindi essere:

la somma di due segnali a 50 dB risulta quindi di 53 dB.
Anatomia dell’apparato uditivo umano
Per capire il motivo per cui la risposta dell’udito umano è diversa a seconda della frequenza del suono, è necessario conoscere la composizione dell’apparato uditivo umano. L’organo dell’udito può essere suddiviso in tre zone specifiche, come si può vedere in figura: l’orecchio esterno, l’orecchio medio e l’orecchio interno.
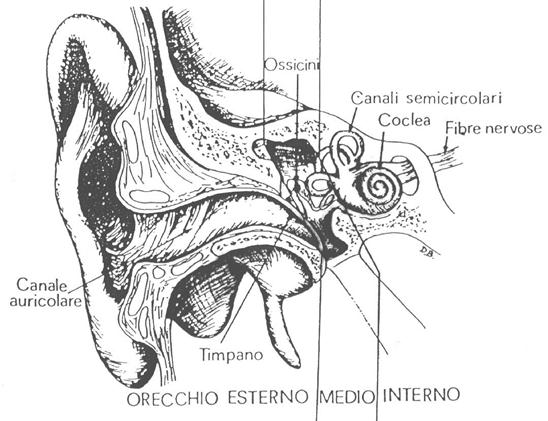
Figura 3: Sezione dell'orecchio umano
L’orecchio esterno comprende il padiglione auricolare, il condotto uditivo e il timpano. Il padiglione auricolare è la parte più esterna del sistema uditivo, la sua funzione è di raccogliere i suoni in modo che attraversino il condotto uditivo. La caratteristica forma a tromba di questo canale provoca il passaggio del suono e una sua leggera amplificazione. Il timpano è una sottile ed elastica membrana, impermeabile ad acqua ed aria, che separa l’orecchio esterno da quello medio, posto alla fine del condotto; quando un suono lo raggiunge ne provoca la vibrazione e questa viene trasmessa alle parti più interne. Il timpano inoltre ha un’impedenza acustica molto piccola, paragonabile a quella dell’aria, in modo che non vi sia una grossa dispersione dell’energia del segnale che trasmette. Questo fatto è aiutato dalla leggera amplificazione provocata dal condotto uditivo: tutto l’orecchio esterno fornisce così la più bassa impedenza acustica possibile e risulta quindi essere un ottimo trasduttore del campo acustico. A causa però della strettezza del condotto stesso il ragionamento appena fatto è valido soprattutto per suoni ad alta frequenza (>1000 Hz), mentre per quelli a basse frequenze la risposta è leggermente minore.
L’orecchio medio è costituito da una cavità interna del cranio, detta cassa timpanica, piena d’aria e contenente una catena di ossicini che hanno il compito di trasmettere la vibrazione del timpano all’orecchio interno; la cassa è collegata alle retrocavità nasali attraverso un condotto detto tromba di Eustachio. Gli ossicini, visibili ingranditi nella figura 4, sono nell’ordine a partire dal timpano: il martello, l’incudine e la staffa. Quest’ultima è appoggiata su un’ulteriore membrana, che separa l’orecchio medio da quello interno, detta finestra ovale.
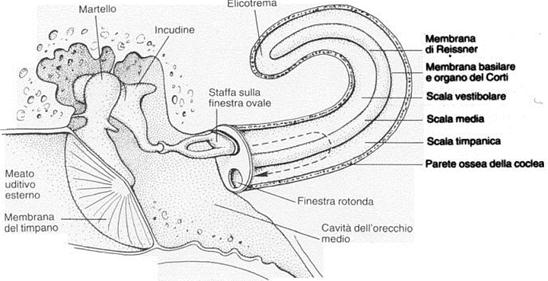
Figura 4: Orecchio medio e Coclea
Poiché l’orecchio interno è riempito da un liquido detto endolinfa caratterizzato da un’impedenza acustica simile a quella dell’acqua, il segnale sonoro trasmesso dall’orecchio medio risulterebbe molto attenuato. Questo problema è risolto dagli ossicini che svolgono la funzione di vere e proprie leve in serie: trasformano il segnale trasmesso dal timpano, caratterizzato da grandi spostamenti ma piccole pressioni, in movimenti della staffa piccoli ma ad alte pressioni. Questo processo non è una vera e propria amplificazione del segnale, esso infatti risulta più intenso ma meno veloce, ma una trasformazione meccanica dell’impedenza.
L’orecchio interno comprende la coclea, il vero e proprio organo dell’udito, e il labirinto, che regola l’equilibrio. La coclea è un tubo a forma di chiocciola suddiviso in due canali (o scale), separati dalla membrana basale: il segnale trasmesso dalla finestra ovale attraversa tutta la coclea, verso il suo centro, lungo il canale vestibolare, in seguito ne esce seguendo in senso opposto il canale timpanico. Durante il passaggio del segnale, la membrana basale si trova sotto sforzo a causa della differenza di pressione presente tra i due canali: questi sforzi vengono registrati dalle cellule cigliate di cui è ricoperta. Queste cellule sono sede di terminazioni nervose e hanno il compito di trasmettere le informazioni al cervello attraverso il nervo acustico.
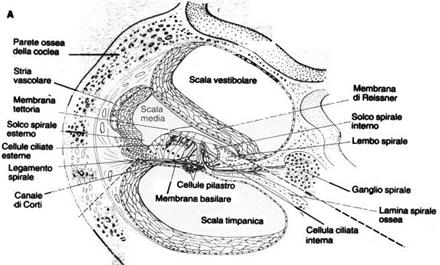
Figura 5: Sezione della coclea
La membrana basale funge inoltre da filtro molto selettivo per quanto riguarda le frequenze percettibili; essa è infatti molto tesa e sottile nelle vicinanze della finestra ovale, mentre diventa sempre più spessa e molle all’avvicinarsi del centro. La tensione della membrana è proporzionale alla frequenza di risonanza a cui lavora: i suoni ad alta frequenza vengono quindi riconosciuti subito, mentre quelli a bassa frequenza devono percorrere tutto il canale vestibolare (ca. 30 mm) prima di essere uditi. Questo fenomeno provoca l’attenuazione dei suoni a basse frequenze e spiega l’andamento delle curve isofoniche del diagramma di Fletcher e Munson.
Un altro aspetto importante che si può osservare nell’apparato uditivo umano risiede nel fatto che il canale di trasmissione del suono è unico e assai limitato: questo fa sì che, quando sono presenti al suo interno troppe informazioni, quelle aventi minore intensità vengono trascurate. Questo accade quando un suono è caratterizzato da diverse componenti, simili in frequenza, ma in cui una prevalga per intensità: la sensazione che riceviamo coincide quindi con la componente più intensa, mentre le altre non sono percepibili. Il fenomeno appena descritto, detto Mascheramento, è alla base delle più moderne tecnologie di compressione dei file audio.
Mascheramento
Come abbiamo visto, la struttura e il funzionamento dell’udito umano comportano la perdita di alcune componenti del suono che percepiamo, a causa del fenomeno chiamato mascheramento. Questo fenomeno può essere visto come unione di due diversi contributi: uno temporale e uno in frequenza.
Il Mascheramento in frequenza è dovuto al fatto che l’udito umano ha una risoluzione limitata in funzione della frequenza: se, ad esempio, consideriamo un tono ad un certo livello e una data gamma di frequenze, la sensazione che ne riceviamo è identica a quella di un tono a livello più basso ma a più ampia banda. In altre parole se l’energia del segnale sonoro è la stessa, a meno di variare banda e intensità, la sensazione che riceviamo è identica. Questo ragionamento è valido all’interno di una certa gamma di frequenze, detta banda critica, caratteristica della frequenza di centro banda; tale comportamento porta a dedurre che il sistema uditivo umano si comporti come un filtro passa-banda. Il grafico in figura 6 mostra le aree di mascheramento per suoni a 60 dB alle varie frequenze: l’unità di misura delle ascisse è il Bark che corrisponde alla larghezza di una banda critica.
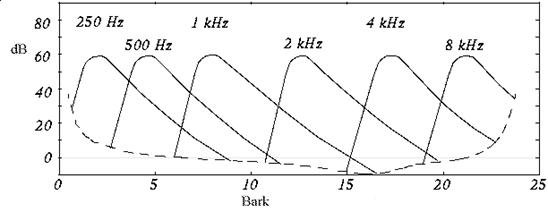
Figura 6: Grafico delle bande critiche
E’ possibile anche scrivere la larghezza delle bande critiche in funzione del centro banda; questi valori, nonostante siano ottenuti adattando valori sperimentali, sono abbastanza attendibili. Per frequenze <500 Hz vale:
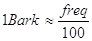
per frequenze >500 Hz vale:

Il Mascheramento temporale è dovuto al fatto che l’udito umano non è in grado di distinguere suoni deboli se ci raggiungono ravvicinati a un tono, simile in frequenza, ma di livello molto alto. In pratica in presenza di un suono forte è necessario che trascorra un dato intervallo di tempo prima di poter distinguere un suono più debole vicino in frequenza. L’effetto di questo fenomeno è ben sintetizzato dal grafico in figura 7:
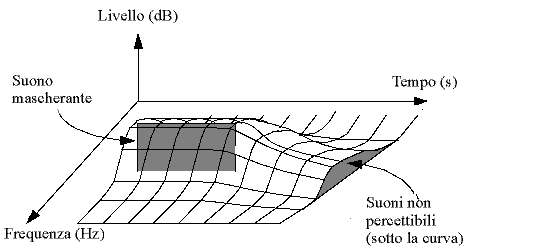
Figura 7: Mascheramento temporale
Il grafico è ottenuto riproducendo un suono con frequenza e livello fissi (il tono mascherante), più un tono di prova di cui si variano frequenza e livello in modo che non venga udito; togliendo il tono mascherante e, dopo un certo ritardo, anche il tono di prova, si regola il ritardo fino al valore più basso per il quale si manifesta il mascheramento.
Compressione sonora
L’archiviazione e la diffusione della musica in formato numerico è una delle più recenti e sviluppate applicazioni dell’acustica psicofisica. Esistono due diverse procedure di archiviazione della musica: la prima, la rappresentazione “senza perdite”, è utilizzata fondamentalmente nei CD audio e si sta espandendo verso una sempre maggiore qualità del segnale immagazzinato; la seconda, la compressione sonora, è utilizzata per la trasmissione via Internet e per l’archiviazione di musica su supporto dati, come ad esempio l’MP3, e si sta espandendo verso una sempre maggiore riduzione del flusso di dati audio senza udibili perdite di qualità. Lo standard CD, che ha una risoluzione dei campioni di 16 bit ed una frequenza di campionamento di 44,1 kHz, genera un flusso di 1,4 Mbit per un secondo di suono stereo: è evidente come risulti difficile immagazzinare o trasmettere segnali sonori di tali dimensioni, da qui la nascita e l’espansione della compressione sonora. Il più diffuso standard di compressione del suono è senza dubbio quello generato dall’MPEG (Moving Picture Experts Group), un gruppo che lavora su standard per la codifica di immagini in movimento e dell’audio.
La compressione sonora si basa sul fenomeno del mascheramento: tutte le componenti di un suono che l’udito umano non riesce a percepire vengono scartate. In questo modo si ha un’effettiva perdita di informazioni, con relativo guadagno nelle dimensioni occupate dal file sonoro, ma la qualità sonora non ne risente, almeno dal punto di vista della sensazione umana. In pratica lo scopo non è quello di restituire il segnale originale intatto, ma piuttosto quello di assicurare che il segnale di uscita suoni uguale al primo per un ascoltatore.
Basandosi su questa filosofia sono stati creati i più importanti standard di compressione, raggiungendo rapporti qualità-dimensioni molto elevati. Le caratteristiche degli standard creati dall’MPEG sono sintetizzate nella seguente tabella:
Livello di compressione |
Standard MPEG |
Caratteristiche |
1 : 4 |
Layer 1 |
384 kb per secondo |
1 : 6 – 1 : 8 |
Layer 2 |
256 – 192 kb per secondo |
1 :10 – 1 : 12 |
Layer 3 (MP3) |
198 – 112 kb per secondo |
E’ interessante osservare come l’algoritmo di compressione alla base dell’MP3 sfrutti appieno i “difetti” del nostro apparato uditivo:
- un banco di filtri scompone il segnale in 32 bande di frequenza, che approssimano le bande critiche;
- seguendo il modello psicoacustico si determina un fattore di mascheramento per ciascuna banda, per fare questo si confrontano i livelli delle bande adiacenti: il fattore di mascheramento coincide con il massimo suono che sarebbe mascherato.
- se il livello di una banda è minore del fattore di mascheramento, l’intera banda non viene codificata;
- si calcola il numero di bit necessario per codificare ciascuna banda, in modo che il rumore introdotto dalla quantizzazione in bit sia minore dell’effetto di mascheramento;
- si crea lo strema audio.
Filtri di ponderazione
Il livello sonoro misurato da uno strumento con una risposta lineare nel campo delle frequenze udibili mal si correla con la risposta soggettiva degli esseri umani allo stesso rumore. Questo perché l’orecchio umano percepisce i suoni secondo una scala logaritmica e in funzione della loro frequenza. Per rimediare a questo fatto si è pensato di introdurre nei misuratori di livello sonoro una ponderazione dei valori misurati in funzione della frequenza, in modo da avvicinarsi alla valutazione non lineare compiuta dagli esseri umani. In particolare, si è notato che prendendo alcune curve isofoniche e ribaltandole si potevano ottenere dei filtri di ponderazione in frequenza fatti in modo che ad una soglia di sensazione più alta corrispondesse una ponderazione più penalizzante.
La curva di ponderazione A è risultata quella in media meglio correlata con la risposta soggettiva umana a rumori generici a larga banda; questo fatto, unito alla facilità di una misurazione fonometrica in dB(A), ha portato all'adozione della curva A in molte norme e leggi nazionali ed internazionali. Quando però il rumore ha forti componenti tonali o è di tipo impulsivo la curva A non da una valutazione adeguata e viene quindi usata la curva C, la cui risposta è misurata in dB(C).

Figura 8: Principali curve di ponderazione
Il filtro A è solitamente usato a valle dei microfoni per misurare i valori efficaci medi e stimare la risposta effettiva dell’orecchio umano. Il filtro C è invece usato per misurare i massimi di picco di suoni forti e impulsivi: in particolare la normativa CEE stabilisce per legge il valore massimo di picco per l’uomo in
Lp,max,peak=130 dB(C)
Perdita uditiva
Con l'incremento dello sviluppo tecnologico, una particolare attenzione merita lo studio dei danni provocati sull'uomo dal rumore, con lo scopo di attuare sistemi preventivi che annullino, o quantomeno riducano al minimo, i suoi effetti. I danni al sistema uditivo possono essere fondamentalmente di due nature diverse: o causati da una breve esposizione a suoni molto intensi (@130 dB), oppure da un’esposizione prolungata a livelli sonori medio-alti (90 – 100 dB).
L’esposizione per poche ore a suoni molto intensi può provocare, a causa dell’elevata sollecitazione meccanica dell’apparato uditivo, un temporaneo malfunzionamento dell’organo intero. Gli effetti sono quindi molteplici, oltre a una temporanea incapacità di percepire altri suoni (mascheramento), possono presentarsi perdita dell’equilibrio, nausea e labirintite. Tali effetti risultano diradarsi nel tempo secondo una legge esponenziale che dipende dall’intensità e dalla durata dell’esposizione; se tali valori superano però una soglia critica si possono presentare danni permanenti: in genere si perde sensibilità per quei suoni caratterizzati da frequenze simili a quello che ha provocato il danno.
Di maggiore interesse sono le patologie provocate dall’esposizione prolungata ad alti livelli sonori: questo è il problema a cui vanno incontro gli operai che lavorano 8 ore al giorno a contatto con attrezzature rumorose. I sintomi provocati sono di tipo soggettivo e variano nel tempo. Nei primi giorni di esposizione si possono presentare acufeni (fischi dovuti a danni al sistema uditivo) e stanchezza generalizzata; col tempo tali disturbi spariscono a causa dell’adattamento. Successivamente, il danno diventa irreversibile e si presentano perdite di sensibilità acustica per suoni di frequenza vicina ai 4000 Hz: si ha quindi uno spostamento della soglia uditiva. Il grafico in figura 9 indica lo spostamento in decibel della soglia uditiva di individui soggetti ad esposizione prolungata per diverse quantità di tempo.
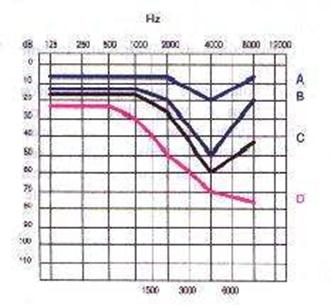
Figura 9: Perdita di udito in ambiente rumoroso
La causa per cui la perdita uditiva maggiore è situata intorno ai 4000Hz, risiede nel fatto che l’organo uditivo umano,in particolare la coclea, è più sensibile a tali frequenze. Tale alta sensibilità è dovuta al fatto che la maggior parte delle componenti della voce umana è caratterizzata da tale frequenza.
In particolare sono le consonanti ad essere caratterizzate dalle alte frequenze, mentre le vocali da quelle più basse (ca. 400Hz). Un individuo affetto quindi da tali lesioni al sistema uditivo, la cosiddetta sordità da rumore (in Italia è presente in ben 2.5 milioni di persone, circa il 5% della popolazione totale), ha quindi la facoltà di sentire la voce umana, ma non riesce a comprendere ciò che gli viene detto.
Fonte: http://www.ramsete.com/dispense00/fossa124322.doc
Apparato uditivo umano
Visita la nostra pagina principale
Apparato uditivo umano
Termini d' uso e privacy