Biochimica del tessuto adiposo
Biochimica del tessuto adiposo
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Le informazioni di medicina e salute contenute nel sito sono di natura generale ed a scopo puramente divulgativo e per questo motivo non possono sostituire in alcun caso il consiglio di un medico (ovvero un soggetto abilitato legalmente alla professione).
Biochimica del tessuto adiposo
Il tessuto adiposo è il tessuto meno ricco di acqua (8-10%), più ricco di trigliceridi (85-90%) e tra i più diffusi (15-20% sul peso totale) dell’organismo.
Più del 95% dei trigliceridi presenti nell’organismo è contenuto nel tessuto adiposo; ciò conferisce quindi al tessuto stesso la funzione di protezione meccanica degli organi interni e di isolamento termico dell’interno organismo.
Inoltre potendo accumulare e degradare trigliceridi ha poi il ruolo fondamentale di riserva di metaboliti energetici.
La capacità di accumulare trigliceridi è pressoché illimitata: le cell di questo tessuto possono andare incontro a ipertrofie da accumulo dell’ordine di migliaia di volte.
È in grado poi di generare calore (termogenesi) contribuendo al mantenimento della temperatura corporea e infine è in grado di secernere sostanze di natura ormonale o ormono-simile.
Esistono 2 tipi di t.adiposo:
- il t.adiposo biancoà di colore giallognolo per la presenza di sostanze pigmentate quali carotenoidi e pigmenti biliari; svolge le funzioni sopra menzionate
- il t.adiposo brunoà di colore scuro per l’abbondanza di mitocondri; nell’adulto è presente in tracce e svolge esclusivamente la funzione termogenica
Tessuto adiposo bianco
Le cell del t.adiposo o adipociti sono una delle forme di differenziazione dei fibroblasti.
Hanno un diametro tra i 20 e i 200µm e può aumentare anche di 20 volte ed il volume di migliaia di volte dietro accentuata sollecitazione nutrizionale.
In condizioni normali hanno una forma rotondeggiante con volume intracell occupato per il 90% da un’unica grossa goccia lipidica.
Il nucleo e i pochi mitocondri sono “schiacciati” contro la membrana plasmatica.
In condizione di digiuno la cell diventa molto + piccola con numerose goccioline lipidiche e molte microinvaginazioni della membrana plasmatica; la morfologia degli adipociti nel digiuno è molto simile a quella dei pre-adipociti.
Gli adipociti sono raccolti in lobuli tenuti insieme da una trama di tessuto connettivale.
Il t.adiposo bianco è più abbondante nella cavità addominale, in corrispondenza del mesentere e attorno ai reni, nelle ghiandole mammarie, nel t.adiposo sottocutaneo nonché tra le fibre del muscolo scheletrico ed intorno al miocardio.
Metabolismo
È incentrato sulla biosintesi e sulla degradazione dei trigliceridi.
Il metabolismo ossidativo, con utilizzo del glucosio e acidi grassi è molto modesto, il consumo di O2 è pure molto basso.
Biosintesi e deposito dei trigliceridi
La disponibilità degli acidi grassi è assicurata da 2 fonti:
- la assunzione dal plasma
- la biosintesi ex novo a partire da acetil-CoA (quantitativamente molto modesto!)
A sua volta l’assunzione dal plasma è condizionata alla loro formazione “in situ” (cioè a livello dello stesso t.adiposo) per l’azione idrolitica esercitata dalla lipoproteina lipasi adiposa sui trigliceridi contenuti nei chilomicroni e nelle VLDL di origine epatica.
La lipoproteina lipasi è secreta verso il letto vascolare e si attacca elettrostaticamente a catene di eparansolfato che sporgono dalla superficie delle cell endoteliari. È a questo livello che avviene l’idrolisi dei trigliceridi e il rilascio degli acidi grassi sia dai chilomicroni che dalle VLDL. Gli acidi grassi fluiscono verso gli adipociti e ne vengono assorbiti verosimilmente con un processo mediato da carrier.
La biosintesi dei trigliceridi richiede la previa attivazione degli acidi grassi ad acil-CoA, operata dalla acil-CoA sintetasi e la presenza di glicerolo-3-fosfato che proviene dal diidrossiaceton-fosfato per riduzione operata dalla glicerolo-3-fosfato deidrogenasi, enzima molto attivo negli adipociti:

Le tappe intermedie della biosintesi sono:
- monoacil-glicerolo-fosfato à acido fosfatidico

- acido fosfatidico à diacilglicerolo
- diacilglicerolo à triacilglicerolo (trigliceride)
catalizzate dalla trigliceride sintetasi (1) e dalla acido fosfatidico fosfatasi.

Il t.adiposo non esprime significative quantità di glicerolo-chinasi: cosicché l’unica sorgente di glicerolo-fosfato è il diidrossiaceton-fosfato e quindi il glucosio.
L’insulina esercita un potente stimolo sul processo di biosintesi dei trigliceridi nel t.adiposo; essa infatti:
- facilita l’ingresso del glucosio negli adipociti, provocando l’inserimento del carrier GLUT4 sulla membrana plasmatica
- stimola la glicolisi mantenendo defosforilata la fruttosio-6-P-chinasi e quindi aumenta la produzione di diidrossiaceton-fosfato
- attiva la lipoproteina lipasi
- attiva gli enzimi della biosintesi ex novo degli acidi grassi e la trigliceride sintetasi
Pertanto tutto il processo di biosintesi è glucosio-dipendente e insulina-dipendente!
Degradazione e mobilizzazione dei trigliceridi
L’idrolisi dei legami esterei dei trigliceridi con liberazione di acidi grassi e glicerolo, richiede l’intervento di 2 enzimi, di cui gli adipociti sono provvisti:
- la trigliceride lipasi, che stacca due resti acilici con formazione del 2-monogliceride
- la monogliceride lipasi che libera il terzo residuo acilico
I prodotti finali sono quindi acidi grassi liberi e glicerolo.
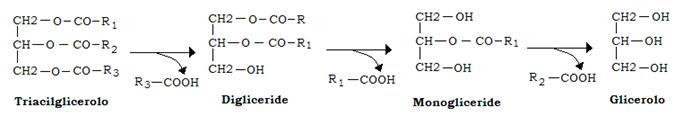
La trigliceride lipasi degli adipociti è anche chiamata “lipasi ormono-sensibile” in quanto la sua attività è sotto stretto controllo ormonale. L’enzima è infatti attivo nella forma fosforilata e inattivo in quella defosforilata.
Pertanto gli ormoni che favoriscono la fosforilazione dipendente dalla presenza di cAMP e quindi dall’innesco del sistema trasduzionale dell’adenilato ciclasi (adrenalina, noradrenalina con i recettori β2; glucagone e ormone della crescita) esercitano un effetto lipolitico, mentre l’insulina, che favorisce la degradazione del cAMP in AMP e attiva le proteine fosfatasi conducendo quindi alla defosforilazione dell’enzima, ne blocca l’azione.
L’effetto antilipolitico dell’insulina è il più precoce e potente dell’ormone!!
A seguito della lipolisi gli acidi grassi che ne risultano passano nel sangue e si ancorano all’albumina che li trasporta alle sedi di utilizzo.
Il glicerolo passa nel sangue e viene utilizzato nel fegato, per la biosintesi di nuovo gliceridi e glucosio.
Funzione secretoria
Gli adipociti producono e secernono numerose proteine le cui azioni si svolgono a distanza, previo ingresso nel torrente circolatorio (proteine ad azione ormonale o ormono-simile) oppure localmente (secreti paracrini).
La meglio nota proteina prodotta e secreta dal t.adiposo è la leptina.
Leptina
È un polipeptide contenente un ponte disolfuro, essenziale per la sua attività. La struttura è simile a quella delle citochine.
È biosintetizzata prevalentemente nel t.adiposo bianco ma anche, se pure in quantità molto minori, nella placenta, nelle ghiandole mammarie, nella mucosa del fundus gastrico e, nei roditori, anche nel muscolo scheletrico.
Tra i principali fattori che regolano l’espressione di leptina hanno particolare rilevanza lo stato nutrizionale e l’attività liposintetica del t.adiposo: maggiore è la liposintesi e quindi la tendenza all’ipertrofia delle cell adipose, maggiore è la produzione ed immissione in circolo della leptina.
Sono state identificate diverse isoforme di recettori della leptina, tutti strutturalmente simili ai recettori delle citochine. Essi possiedono una uguale porzione N-terminale a esposizione extracell. contenente il sito di legame per la leptina, una porzione transmembrana ed una porzione C-terminale intracell che è responsabile dell’innesco del processo di trasduzione del segnale.
I recettori della leptina, a seguito dell’interazione con l’ormone, si dimerizzano rendendosi così disponibili alla fosforilazione su residui di tirosina, catalizzata da una proteina chinasi citosolubile, la JACK.
Il complesso recettoriale così attivato è in grado di fosforilare attivatori trasduzionali di tipo STAT i quali, penetrati nel nucleo sotto forma di omodimeri, attivano per fosforilazione specifici fattori di trascrizione di geni.
Recettori della leptina sono presenti in parecchi nuclei dell’ipotalamo e della base e nel plesso corioideo.
Essi rispondo alla leptina con la secrezione di peptidi ad azione anoressica che riducono il senso della fame e aumentano il senso della sazietà; tra questi figurano il:
- neuropeptide Y (NPY)
- il peptide correlato all’agouti (AcRP)
- la proopiomelanocortina (POMC)
- il trascritto regolato da cocaina e anfetamina (CART)
La leptina stimola anche la secrezione dell’ormone tireotropo con conseguente aumento dell’ormone tiroideo T3; da qui l’incentivazione del metabolismo ossidativo e l’aumento del dispendio energetico.
In conclusione, lo stato di ipertrofia (aumento di dimensione delle cell) del t.adiposo stimola la liberazione di leptina (dal greco “leptos” cioè sottile, magro) la quale, a sua volta, attraverso il controllo dei centri della fame e della sazietà induce una riduzione dell’introduzione di cibo, una intensificazione del dispendio energetico e, di conseguenza, orienta il t.adiposo verso l’ipotrofia.
La leptina circola nel sangue prevalentemente nella forma legata a proteine plasmatiche e passa attraverso la barriera ematoencefalica probabilmente con un processo di transcitosi.
A livello renale passa nel filtrato glomerulare, viene degradata dalle cell tubulari e i frammenti ottenuti vengono escreti con le urine.
Altre proteine secretorie
Il t.adiposo produce proteine implicate nei processi infiammatori:
- l’adipsina (o fattore D del complemento) che attiva per proteolisi alcuni fattore del complemento e ne costituisce la cosiddetta via alternativa
- l’adiponectina che incentiva l’azione dell’insulina e impedisce l’adesione dei alle cell endoteliari esercitando così un’azione sfavorente la formazione della placca aterosclerotica
- alcune citochine, quali il TGF-β che stimola la proliferazione dei precursori degli adipociti e il TNF-α e l’interleuchina 6 che invece tendono a ridurre il trofismo e il differenziamento dei preadipociti in adipociti
- la apolipoproteina E incorporata poi nei chilomicroni, le VLDL e le HDL
- la proteina stimolante l’acilazione che promuove il trasporto degli acidi grassi dal sangue agli adipociti e l’accumulo di trigliceridi
- la resistina ricca di ponti disolfuro che provoca o aumenta la resistenza all’insulina con meccanismo ancora ignoto
- l’inibitore dell’attivatore del plasminogeno PAI.1 agente pro-coagulante e inibitore della fibrinolisi, nonché promotore del rilascio del PDGF
Il t.adiposo contiene inoltre 2 enzimi di notevole importanza nel metabolismo degli ormoni steroidei circolanti:
- 17 β-idrossisteroide ossido riduttasi
- aromatasi citocromo P450 dipendente
La prima converte l’androstenedione in testosterone ormone attivo:
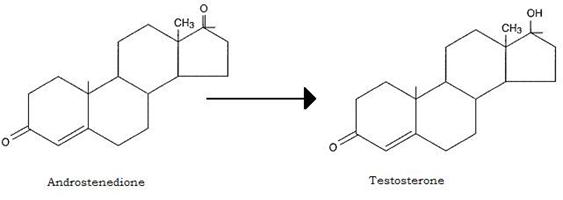
Questa via di produzione del testosterone può, nei soggetti femminili, produrre effetti di mascolinizzazione.
Lo stesso enzima può convertire l’estrone, poco attivo, nell’estradiolo, molto attivo:
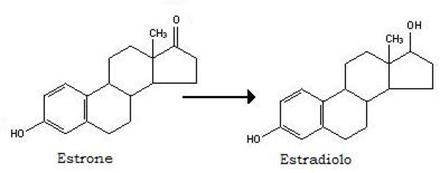
La aromatasi catalizza la aromatizzazione dell’anello A degli androgeni, trasformandoli in estrogeni, per es, il testosterone in estradiolo.
La produzione di tutte queste sostanze aumenta con l’aumentare della massa del t.adiposo!
Termogenesi e tessuto adiposo bruno
La degradazione ossidativa completa dei substrati energetici ha una resa in energia libera, in termini di ATP prodotto, non superiore al 40%; la restante, più rilevante, quota è liberata come calore.
La “normale” produzione di calore che vede implicati soprattutto i tessuti muscolare ed epatico, ma anche il t.adiposo bianco è + che sufficiente per mantenere costante la temperatura corporea.
Pertanto il t.adiposo bruno rimane limitato a piccole isole collocate a livello addominale, del tronco e delle zone sottoscapolare, interscapolare ed ascellare.
Al contrario nella prima infanzia dove il suddetto rapporto è più a favore della superficie di dissipazione del calore, e quindi più calore spontaneamente si dissipa, il t.adiposo bruno è molto più sviluppato e svolge un ruolo non trascurabile nel contribuire a mantenere costante la temperatura corporea.
Metabolismo
Gli adipociti bruni sono più piccoli di quelli bianchi, molto ricchi di mitocondri, con nucleo non schiacciato sotto la membrana plasmatica e con più gocce lipidiche.
La loro peculiarità metabolica consiste nell’esprimere una proteina che disaccoppia la fosforilazione ossidativa dal flusso elettronico della catena respiratoria.
Questa proteina detta anche UCP o termogenina è un canale protonico inserito nella membrana mitocondriale interna.
Con canale aperto i protoni, che nella catena respiratoria funzionante vengono continuamente spostati dalla matrice mitocondriale allo spazio intermembrana, rientrano nella membrana spinti dal gradiente escludendo il passaggio mediato dalla ATP sintetasi.
Si realizza il disaccoppiamento e tutta l’energia del potenziale di ossidoriduzione si libera come calore.
L’UCP è tenuta nella forma aperta da elevate concentrazioni di acidi grassi liberi, condizione che si realizza quando nella cell è in corso intensa lipolisi.
Allorché il flusso lipolitico si sospende e il substrato ossidabile diventa il glucosio, il canale tende a chiudersi, i protoni si impegnano con l’ATP sintetasi e si produce ATP: l’ATP a sua volta mantiene il canale nella forma saldamente chiusa.
Di UCP ne esistono 2 isoforme: una espressa prevalentemente nel grasso bruno e l’altra espressa in modo più ubiquitario.
L’ancoraggio funzionale del t.adiposo bruno alla termogenesi è operato dalle catecolamine, adrenalina e noradrenalina, per le quali gli adipociti bruni dispongono di recettori di tipo β.

Fonte: http://www.medwiki.it/sites/default/files/Biochimica%20del%20tessuto%20adiposo.doc
Sito web da visitare: http://www.medwiki.it
Autore del testo: non indicato nel documento di origine
Parola chiave google : Biochimica del tessuto adiposo tipo file : doc
Visita la nostra pagina principale
Biochimica del tessuto adiposo
Termini d' uso e privacy