Sistema nervoso centrale autonomo e periferico
Sistema nervoso centrale autonomo e periferico
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Le informazioni di medicina e salute contenute nel sito sono di natura generale ed a scopo puramente divulgativo e per questo motivo non possono sostituire in alcun caso il consiglio di un medico (ovvero un soggetto abilitato legalmente alla professione).
Il sistema nervoso
Il sistema nervoso comprende tutti gli organi dell'apparato nervoso che è composto dal sistema nervoso centrale e dal periferico

Il sistema nervoso ha il compito di ricevere gli stimoli esterni e interni all’organismo, di elaborarli e produrre una risposta. Inoltre, si occupa di funzioni psichiche complesse, come la memoria, l’apprendimento e le emozioni.
I segnali esterni sono catturati da cellule specializzate, i recettori, che in molti casi si trovano raggruppate in veri e propri organi di senso. Ogni recettore è specializzato a captare un segnale particolare, luminoso, meccanico, chimico o termico. Se viene stimolato invia dei segnali al sistema nervoso che decide di volta in volta se e come rispondere. Tutto questo è reso possibile dall’attività coordinata di circa 100 miliardi di cellule nervose, che comunicano con una rete di circa 100000 miliardi di fibre di collegamento.
Il sistema nervoso comprende il sistema nervoso centrale e il sistema periferico. Il primo, formato dall'encefalo e dal midollo spinale, interpreta i segnali che provengono sia dall'esterno sia dall'interno del corpo, ed elabora risposte.
Dal sistema nervoso centrale si sviluppa una rete di nervi che raggiunge ogni parte del corpo (sistema nervoso periferico). Il sistema nervoso è formato da miliardi di neuroni collegati insieme che trasportano messaggi in forma di impulsi elettrici che corrono a diverse velocità, da un minimo di 1 km l’ora ad un massimo di 450 come nel caso degli impulsi motori. I neuroni sensitivi trasportano gli impulsi dagli organi di senso, che registrano i cambiamenti relativi all’ambiente interno ed esterno, al sistema nervoso centrale. I neuroni motori portano i segnali dal sistema nervoso centrale verso i muscoli che, in risposta, si contraggono. I neuroni associativi, i più numerosi, si trovano soltanto nel sistema nervoso centrale, dove collegano i neuroni sensitivi e motori formando un complesso centro di elaborazione delle informazioni.
Il sistema nervoso periferico, a sua volta, può essere distinto in sistema nervoso somatico e autonomo o vegetativo, responsabili rispettivamente delle risposte volontarie e involontarie dell’organismo.

La parte superiore dell'encefalo è divisa in due emisferi che dominano uno sull'altro a seconda delle funzioni.

I neuroni
Le cellule più importanti del sistema nervoso, quelle in grado dì ricevere e inviare dei segnali, sono i neuroni. Una delle loro principali caratteristiche infatti è quella di essere provvisti di prolungamenti: i dendriti, che raccolgono i segnali, e di un prolungamento emissario, l'assone, che trasmette i segnali ad altre cellule.
La gran parte dei neuroni ha ramificazioni piuttosto brevi, ma in quelli che raccolgono i segnali dall'esterno e in quelli motori che controllano l'esecuzione del movimento, questi prolungamenti sono molto più lunghi.
L'altra importante caratteristica è lo speciale comportamento delle loro membrane. Quando un dendrite riceve dei segnali da un neurone vicino, viene alterata la carica elettrica della membrana del dendrite, e il cambiamento si diffonde a tutta la cellula fino all'assone, dove costituisce un impulso nervoso. Questo viaggia lungo l'intero assone e alla fine è trasmesso ad altri neuroni.
Le fibre nervose che uniscono il sistema nervoso centrale a tutto il resto del corpo sono costituite da assoni attorno ai quali è avvolta una sostanza bianca, la mielina, che facilita la propagazione dell'impulso nervoso. Più fibre nervose riunite formano i nervi. Dove un assone entra in contatto con un altro neurone o un muscolo, il segnale promuove la liberazione di una sostanza chimica. Questa trasporta il segnale dal neurone trasmittente alle cellule riceventi. Le sostanze usate nella trasmissione sono chiamate neurotrasmettitori, e sinora ne sono state identificate una trentina.

Il cervello
Il cervello è l’organo centrale del sistema nervoso e riceve di continuo segnali che, analizzati, danno luogo a delle risposte di vario tipo. Inoltre, controlla funzioni indispensabili alla sopravvivenza come la temperatura corporea, il battito cardiaco o il respiro. La parte superiore e più ampia è costituita dal cervello propriamente detto, diviso in due metà, gli emisferi cerebrali, che hanno un’organizzazione simmetrica rispetto al corpo. L’emisfero destro controlla la metà sinistra del corpo e quello sinistro la metà destra. Le cellule nervose sono ammassate nello strato più esterno, la corteccia cerebrale, e in alcuni nuclei centrali.
Nell’insieme solo il 25 per cento della corteccia cerebrale ha funzioni definite, per esempio motorie (da cui partono comandi di movimento) o sensitive (dove giungono i segnali dagli organi di senso). Il restante 75 per cento è rappresentato dalle cosiddette aree di associazione, in cui si trovano gran parte dei neuroni che elaborano l’informazione sensoriale prima di decidere cosa fare: è proprio con queste cellule, i cosiddetti interneuroni, che il cervello pensa. Le zone della corteccia che ricevono i segnali sensitivi infatti ne fanno solo un’analisi preliminare. Nel caso di segnali visivi, ad esempio, le aree corticali analizzano i colori o le distanze, ma solo nell’area di associazione visiva un viso o la facciata di un palazzo sono riconosciuti come tali e viene loro attribuito un significato. Le aree di associazione sono poi d’importanza assoluta nelle funzioni mentali superiori come il ragionamento o la creazione artistica.
Il funzionamento del cervello richiede grandi quantità di energia sotto forma di una zucchero semplice, il glucosio. Anche se rappresenta solo il 2 per cento del peso corporeo, il cervello consuma, a riposo, ben il 22 per cento circa di tutte le energie consumate dall’organismo.
Le cellule del talamo raccolgono i segnali nervosi in arrivo e li smistano in parti diverse degli emisferi a seconda dei messaggi che portano.
Gli emisferi cerebrali sono divisi in zone specializzate per funzioni. Per questo si distinguono, ad esempio, una zona motori, una sensitiva, una acustica e una visiva.
Nel corpo calloso passano 300 milioni di fibre nervose che collegano i due emisferi cerebrali
Il bulbo contiene i centri che controllano il battito cardiaco, il respiro e la pressione.
Il pante contiene una rete di cellule, il sistema reticolare attivatore, responsabile dell’alternarsi del sonno e della veglia.
Il cervelletto controlla il tono muscolare e la coordinazione dei movimenti
Nell’ipotalamo si trovano i centri nervosi della fame, della sete, dell’attività sessuale e della temperatura.

I riflessi nervosi
L’uomo è l’animale in cui la superficie del cervello raggiunge le dimensioni più grandi rispetto al corpo. Grazie a questo grande sviluppo le nostre risposte agli stimoli percorrono in genere una strada molto lunga. I segnali arrivano infatti al cervello, dove vengono analizzati in sequenza da molte cellule, per finire eventualmente ai neuroni motori.
Il nostro corpo è però capace anche di risposte nervose involontarie che coinvolgono solo pochi neuroni:i riflessi.
Coi riflessi rispondiamo involontariamente a stimoli di vario genere.
Se possiamo dal buio alla luce,un riflesso nervoso restringe le pupille.
Quando allontaniamo di scatto la mano da un oggetto rovente ,il responsabile è un altro riflesso nervoso.
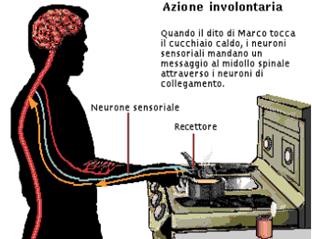
Proprio perché automatici ,i riflessi sono un meccanismo di protezione che non richiede la nostra attenzione. D’altra parte, però forniscono sempre le stesse risposte agli stessi stimoli e, se il nostro comportamento dipendesse solo dai riflessi, saremmo simili a degli automi.
Molti riflessi sono istintivi, sono cioè nostri fin dalla nascita perché codificati geneticamente. Altri invece, i cosiddetti riflessi condizionati, sono il frutto dell’esperienza. La vista di certi piatti, ad esempio, ci fa venire l’acquolina in bocca perché l’esperienza ci ha insegnato che sono buoni.
Il sistema nervoso autonomo
Ciascuno di noi è cosciente soltanto dell’attività del sistema nervoso centrale e periferico, cioè degli stimoli provenienti dall’esterno e delle risposte ad essi date. Ma c’è un’altra sezione del sistema nervoso, della cui attività non ci rendiamo conto, che regola le condizioni dell’ambiente interno del corpo. Si tratta del sistema nervoso autonomo, chiamato anche vegetativo o involontario, costituito da cellule nervoso sparse all’interno del sistema nervoso centrale o in organi distinti, i gangli. Le fibre nervose che escono da questi centri controllano il funzionamento degli organi dell’apparato digerente, cardiocircolatorio, respiratorio, escretore e riproduttivo, e la muscolatura liscia delle arterie.
Il sistema nervoso autonomo è sotto il controllo dell’ipotalamo, che rappresenta il vero centro superiore di tutto il sistema.
L’ippotalamo è collegato, sia pure indirettamente, anche con la corteccia cerebrale: attraverso di esso la vita affettiva e di relazione, che ha luogo nella corteccia, può condizionare l’attività vegetativa.
Organizzazione del sistema nervoso autonomo
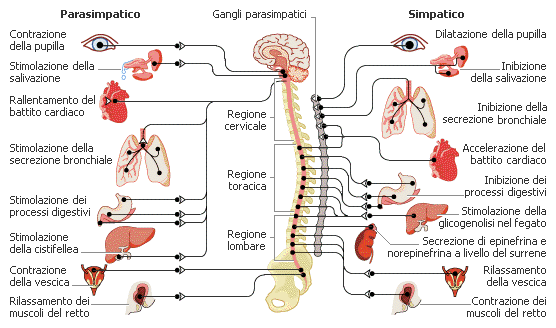
Il sistema nervoso autonomo controlla le funzioni vegetative, ossia le attività involontarie di ghiandole, organi e della muscolatura liscia. Esso è distinto in sistema nervoso autonomo simpatico e sistema parasimpatico. Il sistema simpatico mantiene l'organismo in una condizione di all'erta, preparandolo a una pronta risposta. Il parasimpatico favorisce il risparmio energetico e condizioni di rilassamento dell'organismo. Ciascun nervo del sistema autonomo possiede due neuroni, che portano l'informazione dal sistema nervoso centrale all'organo che esso innerva. La posizione delle sinapsi tra i neuroni è diversa nei due sistemi: nel simpatico essa si trova nei gangli prossimi al midollo spinale; nel parasimpatico è localizzata nei gangli posti in vicinanza o all'interno degli organi.
Le malattie del sistema nervoso
La neurologia si occupa dello studio e della cura dei disturbi del sistema nervoso, mentre la psichiatria si occupa dei disturbi del comportamento
Le malattie del sistema nervoso comprendono alcune malattie genetiche e metaboliche, i disturbi vascolari, le malattie degenerative (tra cui il morbo di Parkinson, il morbo di Alzheimer, le malattie da stress e i tumori, e interessano i neuroni o le altre cellule del tessuto nervoso).
I disturbi vascolari, come l' emorragia cerebrale o altre forme di ictus, sono tra le cause più comuni di paralisi e di altre complicazioni neurologiche. Alcune malattie presentano una particolare distribuzione geografica o per età: ad esempio, la sclerosi multipla è una malattia degenerativa frequente nelle zone temperate, mentre è rara ai tropici.
Il sistema nervoso è soggetto a infezioni provocate da una grande varietà di batteri, parassiti e virus. Ad esempio, il botulismo, la brucellosi, il tetano, la difterite sono patologie causate da batteri, la malattia del sonno è provocata da un parassita, il tripanosoma (trasmesso dalla mosca tse-tse); la meningite è un'infiammazione delle meningi che può essere provocata da molti agenti patogeni diversi, mentre la rabbia è dovuta all'infezione di un unico ceppo virale.
Alcuni virus responsabili di disturbi neurologici colpiscono solo alcune parti del sistema nervoso: il virus responsabile della poliomielite colpisce generalmente il midollo spinale, mentre i virus che provocano l'encefalite attaccano il cervello.
Le infiammazioni del sistema nervoso prendono il nome dalla parte colpita. La mielite è un'infiammazione del midollo spinale, la nevrite è l'infiammazione di un nervo che può essere provocata non solo da un'infezione, ma anche da un avvelenamento, dall'alcolismo o da un trauma. Di solito, i tumori che originano nel sistema nervoso sono formati, a seconda della parte del sistema nervoso colpita, da tessuto meningeo o da cellule di nevroglia (il tessuto di sostegno); altri tipi di tumori possono, invece, diffondersi al sistema nervoso o invaderlo a partire da altre parti del corpo. In alcuni disturbi del sistema nervoso, come la nevralgia, l'emicrania e l'epilessia, non esistono prove dell'esistenza di un danno organico. Un altro disturbo neurologico, la paralisi cerebrale infantile, è associato a un danno subito prima, durante o dopo il parto.
Fonte: http://digilander.libero.it/cfolli/richerche_relazioni/Sistema%20nervoso_Lorenzo.doc
Autore: Lorenzo Grementieri
Sistema nervoso centrale autonomo e periferico
Anatomia e Fisiologia del sistema nervoso
Il sistema nervoso è diviso in due parti: sistema nervoso centrale (SNC) e sistema nervoso periferico (SNP). Il SNC è costituito dall’encefalo e dal midollo spinale, mentre tutto il resto del tessuto nervoso fa parte del SNP che trasporta le informazioni sensitive al SNC e comandi motori ai muscoli e organi.
Il tessuto nervoso possiede 2 tipi di cellule nervose: neuroni e neuroglia.
- Neurone: costituito da un corpo cellulare (soma o pirenoforo) che presenta diversi dendridi con spine dendridiche, il Soma e attaccato ad un lungo assone che termina con una o più terminazioni sinaptiche, attraverso ognuna di esse il neurone comunica con un’altra cellula.
- Neuroglia: si trovano 4 tipi di cellule gliali:
- astrociti- mantenimento barriera ematoencefalica, rete di sostegno,funz. Riparative, controllo ambiente
- oligodendrociti – produzione di mielina
- microglia – ingloba detriti cellulari
- e cellule ependimali – produzione liquor (LSC).
Classificazione strutturale dei Neuroni:
- Neuroni anassonici - SNC
- Neuroni bipolari – visive, olfattive, uditive
- Neuroni unipolare – neuroni sensitivi del SNP, assone mielinico
- Neuroni multipolari – tutti i neuroni che controllano i muscoli scheletrici
Classificazione funzionale dei Neuroni:
- Neuroni sensoriali: quasi tutti Neuroni unipolari, con il loro corpo cellulare al di fuori del SNC, localizzato nei gangli periferici. Formano la porzione afferente del SNP, conducono gli stimoli al SNC. I loro assoni vengono chiamati fibre afferenti si estendono dal recettore sensoriale al midollo spinale o all’encefalo. Raccolgono le informazioni provenienti dall’ambiente esterno (neuroni sensoriali somatici) e interno (neuroni sensoriali viscerali). I recettori possono essere:
- Esterorecettori - che raccolgono le informazioni dall’ambiente esterno.
- Propriocettori – che monitorizzano la posizione e il movimento dei muscoli scheletrici e delle articolazioni.
- Intercettori – che ricevono informazioni dai vari apparati interni, e tattilo dolorifiche, di gusto etc.
- Neuroni motori (motoneuroni): sono neuroni multipolari che formano la componente efferente del sistema nervoso, un motoneurone è in grado di stimolare un tessuto, un organo, o un apparato. Gli assoni si allontanano dal SNC (fibre efferenti) dividendosi nei due sistemi (SNS) sistema nervoso somatico, e (SNA) sistema nervoso autonomo. Il primo comprende tutti i motoneuroni somatici che innervano i muscoli scheletrici. Il soma di questi neuroni si trova nel SNC e i loro assoni si estendono fino alle giunzioni neuromuscolari dei muscoli scheletrici. Il secondo invece comprende tutti i neuroni viscerali che innervano effettori periferici differenti dai muscoli scheletrici. Ci sono 2 gruppi di motoneuroni viscerali, un gruppo con il soma all’interno del SNC e l’altro invece li ha nei gangli periferici. I neuroni del SNC controllano quelli nei gangli che a loro volta controlleranno gli effettori periferici. Gli assoni che si estendono dal SNC ai gangli sono denominate fibre pre-gangliari, quelli che connettono invece le cellule gangliari con gli effettori periferici saranno le fibre post-gangliari.
- Interneuroni: possono trovarsi tra i neuroni sensoriali e i neuroni motori, si trovano esclusivamente nell’encefalo e nel midollo spinale. Hanno la responsabilità di analizzare gli stimoli sensoriali e coordinare gli stimoli motori. Possono essere classificati come Inibitori o Eccitatori.
L’ impulso nervoso
L’eccitabilità è la capacità di una membrana cellulare di condurre impulsivi di tipo elettrico (potenziale d’azione) quando viene stimolata ad un livello soglia. Raggiunto questo livello cambia la permeabilità della membrana agli ioni sodio e potassio, i movimenti di ioni che ne derivano producono un improvviso cambiamento del potenziale transmembranario (potenziale d’azione). Se questo potenziale d’azione viaggia lungo l’assone viene definito impulso nervoso. La sua velocità dipendono dal diametro dell’assone e dalla presenza della guaina mielinica.(nodi di ranvier).
La comunicazione sinaptica:
Può essere chimica o elettrica:
Le più numerose sono quelle chimiche, dove un neurotrasmettitore rilasciato a livello della membrana pre-sinaptica di un bottone sinaptico lega un recettore sulla membrana post-sinaptica e innesca un cambiamento di potenziale di membrana della cellula recettiva. La comunicazione può avere solo una direzione.
(acetilcolina neurotrasmettitore Ach ). Alcuni neurotrasmettitore avranno effetti eccitatori altri inibitori. L’attività di un neurone recettoriale sarà data dalla somma di tutti gli impulsi sia inibitori che eccitatori.
Le sinapsi elettriche invece si trovano tra i neuroni del SNC-SNP ma sono piuttosto rare. Le due membrane pre e post sinaptiche sono così a stretto contatto da funzionare come se fossero una membrana unica e il messaggio quindi continuerà nella sua corsa senza interruzioni.
Organizzazione e funzione neuronale
Gl’interneuroni sono organizzati in gruppi chiamati appunto gruppi neuronali.
Un gruppo neuronale è un’insieme di neuroni connessi tra di loro con specifiche funzioni, e sono definiti in base alle funzioni piuttosto che su base anatomica, così avremo un gruppo che potrà essere diffuso coinvolgendo neuroni in diverse regioni dell’encefalo, oppure localizzato in una specifica area come un Nucleo.
Il tipo di interazioni tra neuroni caratterizza la funzione di quel gruppo neuronale.
- Divergenza- propagazione di un’informazione da un neurone verso più neuroni
- Convergenza- opposto di sopra così si potrà volontariamente o involontariamente muovere ad esempio un muscolo.
- Processamento seriale –
- Parallelo – la possibilità di portare l’informazione allo stesso tempo in più punti
- Riverberazione – la continuità dell’impulso.
Organizzazione anatomica SN
La funzione del sistema nervoso dipende dalle interazioni tra neuroni a livello dei gruppi neuronali, le più complesse interazioni avvengono al livello del SNC. L’informazione afferente sensoriale e quella efferente motoria sono trasportati dal SNP. Gl’assoni e i peirenofori sono disposti con un ordine ben preciso.
Nel SNP
- i Pirenofori dei neuroni sensitivi e dei neuroni motori si trovano nei gangli
- Un gruppo di assoni costituisce i Nervi; i nervi spinali sono connessi al midollo spinale e i nervi cranici all’encefalo.
Nel SNC
- Un insieme di Pirenofori con una funzione comune è chiamato centro. Un centro con un confine anatomico preciso è detto Nucleo. Porzioni di superfice celebrale sono ricoperte di sostanza grigia detta corteccia celebrale. Il termine centri superiori si riferisce ai centri di integrazione, ai nuclei, e alle aree corticali più complesse dell’encefalo
- La sostanza bianca del SNC contiene fasci di assoni che possiedono origini, destinazioni e funzioni comuni. Questi fasci sono detti tratti. Tratti nel midollo spinale formano ampi gruppi, detti colonne.
- I centri e i tratti che collegano l’encefalo con il resto del corpo sono dette vie. Ad esempio, vie sensitive (ascendenti) trasportano informazioni dai recettori periferici ai centri di processamento dell’encefalo, e le vie motorie (discendenti) partono dai centri del SNC che regolano il controllo motorio e finiscono a livello degl’organi effettori che essi controllano.
Anatomia dissettoria del midollo spinale
All’interno del midollo sezionato possiamo trovare una zona centrale a forma di H formata da sostanza grigia, costituita dai corpi cellulari (pirenofori) dei neuroni e da cellule gliali. Le proiezioni della sostanza grigia in avanti e indietro vengono chiamate corna anteriori le prime e corna posteriori le seconde. La sostanza bianca è situata perifericamente e contiene un gran numero di assoni mielinici e amielinici organizzati i tratti e colonne.
Organizzazione sostanza grigia:
I pirenofori dei neuroni della sostanza Grigia del midollo spinale sono organizzati in gruppi (nuclei) con specifiche funzioni. Nuclei sensoriali dorsali: ricevono ed integrano informazioni sensoriali dai recettori periferici (tattili). Nuclei motori ventrali indirizzano comandi motori agli effettori periferici, come i muscoli scheletrici. Possono estendersi per una considerevole lunghezza.
Le corna grigie posteriori contengono nuclei sensoriali somatici, le corna grigie anteriori presidiano al controllo motorio somatico. Le corna grigie laterali contengono neuroni motori viscerali.
Organizzazione sostanza bianca:
Può essere suddivisa in regioni (colonne), colonne bianche posteriori, colonne bianche anteriori. Ogni colonna contiene tratti o fasci che trasportano sia informazioni sensitive che comandi motori e gli assoni sono relativamente uguali per caratteristiche. I fasci ascendenti portano informazioni sensoriali verso l’encefalo e quelli discendenti conducono le informazioni motorie al midollo spinale.
I Riflessi
Risposta motoria involontaria immediata ad uno stimolo specifico. Il percorso di un singolo riflesso prende il nome di Arco Riflesso: inizia al livello di un recettore e termina in periferia su di un organo effettore. Le 5 fasi dell’arco riflesso
- Arrivo di uno stimolo e attivazione di un recettore
- Trasporto dell’informazione al SNC
- Elaborazione dell’informazione
- Attivazione di neurone motorio
- Risposta di un effettore periferico.
Classificazione dei riflessi:
Possono essere suddivisi a seconda:
- del loro sviluppo innati o acquisiti
- del luogo in cui avviene l’elaborazione spinali o cranici
- della natura della risposta motoria che ne deriva somatica,viscerale o autonoma
- della complessità dei circuiti neuronali coinvolti monosinaptici la trasmissione attraverso una sinapsi chimica coinvolge sempre un ritardo sinaptico, ma con una sola sinapsi il ritardo tra stimolo e risposta è minimo.(arco riflesso). I riflessi polisinaptici invece hanno un ritardo più lungo tra stimolo e risposta. Possono produrre risposte più complicate poiché gli interneuroni possono controllare diversi gruppi muscolari.
- Riflesso monosinaptico spinale da stiramento: consiste in una regolazione automatica della lunghezza del muscolo scheletrico. Lo stimolo allo stiramento di un muscolo rilassato attiva un neurone sensoriale che innesca la contrazione di quel muscolo.
Organizzazione dell’ Encefalo
L’encefalo adulto può essere suddiviso in 6 principali regioni:
- cervello
- diencefalo
- mesencefalo
- ponte
- cervelletto
- bulbo (midollo allungato)
Cervello
Suddiviso in due ampi emisferi cerebrali, separati da una scissura longitudinale:(Coscienza, funzioni intellettuali, memorizzazione, ricordo e complesse attività motorie si originano nel cervello. Ogni emisfero può essere suddiviso in lobi ben delineati nei loro confini anatomici che prendono il nome dall’osso cranico sovrastante. Ogni lobo contiene regioni funzionali i cui confini sono cosi delineati
- lobo frontale –corteccia motoria primaria- controllo movimenti volontari. I neuroni della corteccia motoria primaria controllano la muscolatura volontaria, esercitando la loro azione sui neuroni presenti a livello del tronco cerebrale e del midollo spinale. I neuroni della corteccia motoria primaria prendono il nome di NEURONI PIRAMIDALI e la via che svolge il controllo della muscolatura volontaria è detta VIA CORTICOSPINALE o SISTEMA PIRAMIDALE
- lobo parietale –corteccia sensitiva primaria- percezione conscia del tatto, pressione, vibrazione, temperatura, gusto. L’informazione che raggiunge questi neuroni è conscia, ma allo stesso tempo, tramite rami collaterali l’informazione sensoriale può raggiungere i nuclei cerebrali e altri centri nervosi in questa maniera l’informazione sensoriale sarà controllata sia a livello conscio che a livello inconscio.
- lobo occipitale –corteccia visiva- percezione coscia degli stimoli visivi
- lobo temporale –corteccia uditiva e olfattoria- percezione conciai degli stimoli uditivi e olfattivi
- tutti i lobi – aree associative- integrazione ed elaborazione dei dati sensitivi, elaborazione ed inizio delle attività motorie.
- area associativa motoria somatica –corteccia premotoria- è responsabile per la coordinazione delle risposte motorie apprese
- area associativa sensoriale somatica – riceve input dalla corteccia sensitiva primaria, dal talamo e altre regioni encefaliche, questa regione integra e interpreta sensazioni concernenti la dimensioni e la forma degli oggetti
I Centri integrativi ricevono e processano le informazioni da varie aree associative, essi controllano attività motorie complesse e svolgono complicate funzioni analitiche. (la corteccia prefrontale del lobo frontale è quella deputata al prevedere la conseguenza ad una eventuale risposta.)
Diencefalo
Situato subito sotto il cervello il diencefalo viene suddiviso in tre regioni
- Epitalamo – detto tetto del diencefalo comprende la ghiandola pineale (Epifisi) che secerne melatonina regolatore dei cicli circadiani.
- Talami – dx e sn che costituiscono le pareti del diencefalo centro di processamento di informazioni sensitive. I nuclei talamici sono centri di controllo di informazioni sensitive e motorie. Le informazioni sensitive ascendenti sono processate nei nuclei talamici prima di essere trasportate nel cervello o al tronco cerebrale. Esso agisce come un filtro, permettendo il passaggio solo ad una parte delle informazioni che vi giungono. In più è un’importante stazione di controllo per le attività motorie volontarie e involontarie. Il talamo a 5 nuclei principali.
- Nuclei del gruppo anteriore fanno parte del sistema limbico e svolgono un ruolo importante nelle emozioni, nella memoria, e nell’apprendimento.
- Nuclei del gruppo mediale mantenimento della coscienza negli stati emozionali, collegano le informazioni sensoriali di altri parti del talamo ai lobi frontali.
- Nuclei del gruppo ventrale trasmettono le informazioni riguardanti comandi motori somatici dai nuclei della base e dal cervelletto alla corteccia motoria primaria e alla premotoria. Pianificano il movimento e aiutano a metterlo in pratica.
- Nuclei del gruppo posteriore comprendono il pulvinar e i nuclei genicolati. Il pulvinar integra informazioni sensoriali per proiettarle alle aree associative della corteccia. Il nucleo genicolato laterale di ogni talamo riceve informazioni visive dalla retina per mezzo dei tratti ottici, che si proiettano nella corteccia visiva e scendono al mesencefalo. Il nucleo genicolato mediale invece riceve le informazioni uditive dirette alla corteccia uditiva
- Nuclei laterali circuiti emozionali e sensitive.
- Ipotalamo – pavimento del diencefalo, centro del controllo viscerale, controllo delle emozioni e produzione ormonale esso è collegato alla ghiandola pituitaria (Ipofisi)
- Controllo subconscio della muscolatura scheletrica (collera, piacere, dolore).
- Controllo delle funzioni autonome centri ipotalamici correggono e coordinano le attività di centri autonomi in altre parti del tronco cerebrale, correlate con la regolazione della frequenza cardiaca, pressione ematica, respiratoria, digestiva.
- Coordinazione delle attività del sistema nervoso ed endocrino inibizione e stimolazione dell’ipofisi.
- Secrezione di 2 ormoni: ormone antidiuretico che riduce l’eliminazione dell’acqua a livello renale, e l’ormone ossitocina muscolatura liscia uterina, prostata e ghiandola mammaria.
- Produzione delle emozioni e condotta comportamentale.
- Coordinamento tra le funzioni volontarie e quelle autonome.
- Regolazione della temperatura corporea
- Controllo del ritmo circadiano
Mesencefalo
I nuclei presenti nel Mesencefalo processano informazioni visive e uditive e generano risposte motorie somatiche involontarie a questi stimoli + alcuni centri del mantenimento della coscienza .
Ponte
Subito sotto il mesencefalo contiene Nuclei coinvolti nel controllo motorio viscerale e somatico, connette il cervelletto al tronco cerebrale,
Cervelletto
Coordinare e correggerei automaticamente le attività motorie sulla base di informazioni sensoriali e del ricordo di movimenti già eseguiti.
Bulbo
E’ il punto di connessione tra il midollo spinale e l’encefalo, la parte superiore ha un tetto la caudale assomiglia al midollo spinale. Le informazioni sensoriali vengono indirizzate al talamo e agli altri centri del tronco cerebrale; inoltre contiene i principali centri per l’integrazione delle funzioni autonome (frequenza cardiaca, pressione ematica, attività digestiva).
Vascolarizzazione dell’encefalo
I neuroni hanno un elevata richiesta di energia, essendo privi sia di riserve energetiche sotto forma di lipidi e carboidrati, che di mioglobina come forma di deposito di ossigeno. Il sangue arterioso raggiunge l’encefalo attraverso le arterie carotidi interne e le arterie vertebrali. Il sangue venoso invece lascia l’encefalo attraverso le vene giugulari interne.
La sostanza bianca è coperta dalla sostanza grigia della corteccia cerebrale. Essa contiene fibre mieliniche che formano fasci che si estendono da un’area corticale ad un’altra o che connettono aree della corteccia con altre regioni del encefalo.
Ci sono tre tipi di questi fasci:
- Fibre associative interconnettono porzioni di corteccia cerebrale nello stesso emisfero cerebrale,
- Fibre commessurali permettono la comunicazione tra i due emisferi,i più importanti corpo calloso, commessura anteriore.
- Fibre di proiezione collegano la corteccia cerebrale al diencefalo al tronco encefalico e al midollo spinale. Tutti gli assoni ascendenti o discendenti devono stazionare nel diencefalo nel loro percorso verso le aree sensoriali o motorie o associative della corteccia. Questo insieme di fasci viene denominato capsula interna.
Nuclei Celebrali - Gangli della base- Nuclei basali
Sono raggruppamenti di sostanza grigia all’interno degli emisferi cerebrali. Tali nuclei si trovano in ogni emisfero al di sotto dei ventricoli laterali, e sono circondati da sostanza bianca e tra essi o intorno ad essi vi sono fibre di proiezione o commessurali.
Corpo striato A+B
Nucleo Caudato – possiede una gran testa e un’esile coda che curva seguendo la curva del ventricolo all’apice della coda vi è un nucleo (amigdala)
Claustro
- Nucleo Lenticolare
- Putamen
- Globus Pallidus
I nuclei celebrali sono coinvolti con il controllo subconscio del tono muscolare (definizione: grado residuo di contrazione leggera e continua che persiste nel muscolo a riposo è importante per la funzione della postura.) e la coordinazione dei movimenti appresi. Questi nuclei non iniziano particolari movimenti. Ma una volta che il movimento sta per partire essi assicurano il ritmo generale del movimento.
Nucleo caudato – quando camminiamo è il caudato con il Putamen che controllano l’alternanza dei movimenti,delle braccia e delle gambe, che intercorrono tra il momento in cui si è deciso di iniziare a camminare e quello in cui si decide di farlo. (durante la camminata)
Claustro – processi inconsci di informazione visiva
Amigdala -
Globus Pallidus – controlla e corregge il tono muscolare per preparare il corpo ad un movimento volontario. (I movimenti involontari di tutto un braccio per poter poi prendere una penna che è il movimento volontario.)
Sistema limbico – Comprende quei nuclei e quei fasci che si trovano tra cervello e diencefalo. Le sue funzioni sono
Controllo degli stati emozionali e conduzioni dei comportamenti correlati
- Collegamento delle funzioni inconsce ed autonome di parti dell’encefalo
- Archiviazione della memoria
Amigdala è un centro di integrazione tra il sistema limbico, il cervello e vari sistemi sensoriali.
Lobo limbico giri e strutture più profonde adiacenti l’encefalo.
- il giro del cingolo
- corpo calloso
- giro dentato
- giro paraippocampale
- ippocampo – apprendimento e memoria a lungo termine
- fornice – sostanza bianca che collega l’ippocampo con l’ipotalamo,
Molte di queste fibre terminano nei corpi mammillari (nuclei presenti nel pavimento dell’ipotalamo) che contengono nuclei motori che controllano i movimenti riflessi associati con il mangiare, come il masticare, e l’inghiottire.
La formazione reticolare è una rete di nuclei interconnessi che ha il quartier generale nel mesencefalo.
Fonte: http://crocelodigiana.it/files/medicina_-_anatomia_e_fisiologia_del_sistema_nervo.doc
Autore del testo: non indicato nel documento di origine
IL SISTEMA NERVOSO
I livelli citosolici di calcio sono piuttosto bassi, circa 100 nM e quindi, essendo la concentrazione nel reticolo più alta, l’apertura dei canali per il calcio induce la fuoriuscita di calcio. All’esterno della cellula la concentrazione del calcio è poco maggiore di 1 mM molto maggiore di 100 nM del citosol. Dato che i canali del calcio sono chiusi, il calcio extracellulare non può fluire all’interno della cellula, ma nel caso in cui i canali si aprono, il calcio può entrare. Ci sono quindi questi due modi per far entrare il calcio all’interno della cellula (citosol). L’entrata del calcio è importante per il rilascio dei neurotrasmettitori dei neuroni. L’aumento del calcio porta all’attivazione di alcune proteine chinasi C. Alcune di queste sono attivate contemporaneamente dal calcio e dal DAG. La calmodulina è una proteina con 4 siti di legame per il calcio. E’ una sorta di sensore per il calcio. L’attività di molte proteine viene modulata dai livelli citosolici di calcio, per interazione diretta o per attivazione indiretta. Il calcio satura i legame della calmodulina, e calcio più calmodulina, si legano ad una certa proteina modulandola. La calmodulina è una proteina calcio dipendente che viene attivata dal calcio. Alcuni trasportatori possono essere attivati dalla calcio-calmodulina. La concentrazione citosolica del calcio è tenuta bassa grazie a trasportatori attivi situati sulla MP e sul reticolo endoplasmatico. La cellula tende a stabilire i valori iniziali sia sequestrando il calcio nel reticolo endoplasmatico, sia estrudendolo nello spazio extracellulare. Nella maggior parte dei tipi cellulari le pompe del calcio estrudono contro gradiente il calcio e sono attivate proprio dalla calmodulina. Questo avviene dopo l’interazione del calcio con la calmodulina. I recettori eptaelica sono una componente importante del sistema di trasduzione del segnale. Circa il 60% di tutti i tipi recettoriali che mediano la trasmissione del segnale, sono recettori accoppiati a proteine G. Sono tipi recettoriali molto diffusi che legano una grande varietà di ligandi come l’acetilcolina e le catecolamine. Adrenalina e noradrenalina (che dovrebbero essere catecolamine) si legano a recettori adrenergici di cui ne esistono vari sottotipi con diversi effettori. Incontremo questi recettori quando parleremo della modulazione dell’attività cardiaca da parte del sistema parasimpatico. L’acetilcolina può legarsi invece a recettori eptaelica muscarinici. Un ligando non necessariamente attiva un solo tipo di recettore, ma può legarsi a più recettori specifici strutturalmente e funzionalmente diversi. Per esempio l’acetilcolina è un neurotrasmettitore importante che attiva i recettori muscarinici accoppiati a proteine G. L’acetilcolina è però anche un ligando di un importante canale ionico (recettore canale) che è un recettore nicotinico. L’acetilcolina può quindi legarsi, o a un recettore muscarinico, o attivare vari tipi di proteine G, o legarsi ad un canale ionico nicotinico innescando una serie di eventi elettrici. Il recettore nicotinico produce una risposta molto rapida. Una volta che il ligando si lega al canale cationico (ioni +), l’effetto prodotto dall’acetilcolina sull’apertura del canale è un fenomeno molto rapido dell’ordine di grandezza del millisecondo. La risposta dei recettori muscarinici è invece una risposta più lenta. Non pochi neurotrasmettitori utilizzano queste due vie. Essi hanno sia recettori canale detti ionotropi, che recettori accoppiati a proteine G detti metabotropi.
(thanks to Marco)
Le cellule nervose, neuroni, trasportano segnali elettrici rapidamente a grande distanza. I neuroni si collegano fra loro per formare archi riflessi.. Nella maggior parte dei riflessi i neuroni comunicano mediante segnali chimici, neurotrasmettitori, attraverso una piccola soluzione di continuo, la sinapsi. In alcuni altri riflessi i neuroni sono legati da giunzioni comunicanti, il che permette ai segnali elettrici di passare direttamente tra le cellule.
I recettori sensitivi tengono sotto controllo le condizioni degli ambienti esterno ed interno e inviano informazione lungo neuroni sensitivi, o afferenti, all’SNC. Il sistema nervoso centrale (SNC) è formato da encefalo e midollo spinale. I neuroni dell’SNC integrano le informazioni in entrata e decidono se è necessaria una risposta. I segnali che dirigono la risposta appropriata vengono inviati in periferia alle cellula effettrici da neuroni efferenti. I neuroni afferenti e quelli efferenti formano il sistema nervoso periferico. I neuroni efferenti dell’ SNP sono suddivisi nella sezione motoria somatica che controlla i muscoli scheletrici, e nella sezione autonoma che governa muscolatura liscia, miocardio e ghiandole. I neuroni autonomi si dividono nei sistemi simpatico e parasimpatico.
Il sistema nevoso è composto da cellule di supporto, le glia, e le cellule nervose vere, i neuroni. Il neurone è l’unità funzionale del sistema nervoso ed ha lunghe appendici che si allontanano dal corpo cellulare, dendriti oppure assoni.
I neuroni si possono classificare secondo la struttura o secondo la funzione. Dal punto di vista strutturale, si classificano in base al numero di processi che si dipartono dal corpo cellulare. In base alla loro funzione si dividono in neuroni (afferenti) sensitivi, interneuroni e neuroni efferenti.
I neuroni sensitivi portano informazioni dai recettori sensitivi periferici all’SNC. I neuroni sensitivi somatici hanno corpi cellulari vicini all’SNC, ed hanno un unico processo, un assone, che si biforca poco dopo l’insorgenza per dare origine a due rami: uno va alla periferia , e in realtà funziona da dendrite, cioè raccogli gli stimoli, mentre l’altro va all’SNC.
I neuroni che sono situati internamente entro l’SNC sono detti interneuroni, hanno ramificazioni molto complesse, che formano sinapsi con molti altri neuroni.
I neuroni efferenti sono simili al classico modello di neurone.
Il corpo cellulare, soma, di un neurone assomiglia alla cellula tipica con nucleo e organuli che dirigono l’attività cellulare. Un ampio citoscheletro si estende entro l’assone e i dendrite. La posizione del soma varia nei diversi tipi di neurone, ma anella maggior parte dei casi è piccolo.
I dendriti sono ramificazioni sottili la cui principale funzione è ricevere segnali in entrata. I neuroni più semplici possono avere un dendrite singolo, le cellule cerebrali hanno invece molti dendriti a ramificazione molto complessi. I segnali che raggiungono i dendriti sono trasferiti a una regione di integrazione nel neurone.
La maggior parte dei neuroni ha un solo assone, una proiezione del corpo cellulare che ha origine da una speciale regione del corpo cellulare, la cresta assonale. Un nervo è un insieme di assoni che trasportano informazioni tra l’SNC e i recettori o cellule bersaglio. Gli assoni trasmettono segnali elettrici dal centro integrativo del neurone alla periferia. All’estremità distale dell’assone, il segnale elettrico viene trasdotto in messaggio chimico con la secrezione di una molecole neurocrina, neurotrasmettitore. Gli assoni possono ramificarsi in vari punti lungo tutta la loro lunghezza, formando collaterali, ogni collaterale termina in un rigonfiamento, il terminale assonale. La regione in cui il terminale assonale incontra la sua cellula bersaglio è detta sinapsi. Il neurone che rilascia il segnale alla sinapsi è detto presinaptico, la cellula che riceve il segnale è detta postsinaptica.
Il trasporto assonale può essere:
- lento: muove materiale dal soma al terminale assonale, trasporta componenti che non sono consumati rapidamente dalla cellula, come enzimi e proteine del citoscheletro
- veloce: è bidirezionale; il trasporto anterogrado (in avanti) trasporta vescicole sinaptiche e secretorie dal soma al terminale assonale, mentre quello retrogrado (all’indietro) trasporta vecchie componenti di membrana dal terminale assonale al soma perché siano riciclate.
Le cellule gliali sono cellule di supporto, non partecipano direttamente alla trasmissione dei segnali elettrici, comunicano con i neuroni tra di loro mediante segnali elettrici e chimici. La glia fornisce sostegno fisico per i neuroni.
I nervi e i muscoli sono tessuti eccitabli, cioè in grado di generare e propagare segnali elettrici. Due sono i tipi di segnali elettrici che sono creati nei neuroni.
- potenziali graduati: che restano a brave distanza dal punto di origine
- potenziali di azione: che viaggiano molto rapidamente sulla lunga distanza.
Tutte le cellule hanno una differenza di potenziale di membrana di riposo. Questo potenziale di membrana è la differenza di potenziale che risulta da una ineguale distribuzione di ioni sui due versanti della membrana.
La cellula a riposo fa passare una quantità molto scarsa di Na+. Di conseguenza il potassio che può uscire dalla cellula seguendo il gradiente, è lo ione che più contribuisce al potenziale di membrana a riposo.
Sebbene l’Na contribuisca in misura minima al potenziale di membrana a riposo, gioca un ruolo chiave nella generazione di segnali elettrici nei tessuti eccitabili A riposo, la membrana del neurone è quasi impermeabile al Na. Se la membrana aumenta improvvisamente la propria permeabilità all’Na, questo attraverserà la membrana. Essendo l’Na più concentrato nel liquido extracellulare, il gradiente di concentrazione favorisce la sua diffusione verso l’interno della cellula. L’aggiunta di ioni positivi nella cellula ne depolarizza la membrana e genera un segnale elettrico.
Il movimento di ioni attraverso la membrana può anche iperpolarizzare la cellula: se la membrana diventa più permeabile al K, una carica positiva è perduta dalla cellula, che diventa più negativa.
Perché una cellula cambi la sua permeabilità ionica, devono esserci nella membrana dei canali ionici, che si aprono e si chiudono. I neuroni possiedono numerosi canali ionici a cancello, che variano tra gli stati di apertura e chiusura in base alle condizioni intracellulari.
La permeabilità al sodio del neurone aumenta quando i suoi canali si aprono in risposta a segnali esterni o interni. I canali per l’Na regolati meccanicamente si trovano i neuroni sensitivi e si aprono in risposta a stimoli fisici. I canali regolati chimicamente rispondono a diversi ligandi che si trovano negli ambienti interno ed esterno, e a neurotrasmettori. I canali per l’Na voltaggio dipendenti giocano un ruolo importante nella conduzione dei segnali elettrici lungo l’assone.
Il sodio non è il solo ione che attraversa le membrane neuronali mediante canali cancello. I canali per il K voltaggio dipendenti si aprono in risposta alla depolarizzazione della membrana, permettendo al K di fluire dal citoplasma al liquido extracellulare lungo il proprio gradiente di concentrazione: lo stesso fanno i canali per il Ca voltaggio dipendenti, quando si aprono il calcio entra nella cellula. Gli ioni calcio sono il segnale per l’avvio dell’esocitosi del neurotrasmettitore nella sinapsi. I canali per il Cl- regolati chimicamente si aprono in seguito al legame con vari neurotrasmettitori: questi canali permettono agli ioni calcio di entrare nella cellula lungo gradiente di concentrazione, iperpolarizzandola.
La trasmissione dei segnali elettrici nei neuroni è il risultato di variazioni del potenziale di membrana. Il movimento netto di cariche elettriche a cavallo della membranadepolarizza o iperpoplarizza la cellula, generando un segnale elettrico.
Due sono i tipi di segnali elettrici neuronali:
- potenziali graduati: sono segnali ad intensità variabile che si smorzano viaggiando lungo la cellula
- potenziali di azione: possono viaggiare su lunghe distanze lungo il neurone senza perdere di intensità.
I potenziali graduati sono depolarizzazioni o iperpolarizzazioni che si verificano nei dendriti o nel soma. La loro ampiezza (forza) è direttamente proporzionale alla forza dell’evento che li ha scatenati: uno stimolo intenso genererà un potenziale graduato forte, un piccolo stimolo provocherà un potenziale graduato debole.
Cominciano dal punto in cui gli ioni entrano dal liquido extracellulare.
Supponiamo che un neurotrasmettitore si combini con recettori situati su un dendrite, aprendo i canali per l’Na. Le cariche elettriche portate dagli ioni sodio si diffondono come un’onda di depolarizzazione nel citoplasma. L’onda di depolarizzazione che si diffonde nella cellula è nota come flusso locale di corrente. La forza della depolarizzazione iniziale in un potenziale graduato è determinata dalla quantità di carica che entra nella cellula. Se si aprono i canali per l’Na , entrano più ioni e il potenziale graduato ha un’ampiezza iniziale maggiore. Maggiore è l’ampiezza iniziale, più lontano diffonderà il potenziale graduato prima di estinguersi.
I potenziali graduali possono essere iperpolarizzati o depolarizzanti e si verificano tipicamente nel soma e nei dendriti. Nei neuroni interni all’SNC e in quelli efferenti, i potenziali graduati si verificano quando segnali chimici provenienti da altri neuroni aprono i canali ionici regolati chimicamente.
I potenziali graduati viaggiano lungo i neuroni fino a raggiungere la zona trigger. Nei neuroni efferenti, la zona trigger si trova presso la cresta assonale e nel segmento iniziale dell’assone. Nei neuroni sensitivi la zona trigger è immediatamente adiacente al recettore, dove i dendriti si uniscono all’assone.
Se i potenziali graduati che raggiungono la zona trigger depolarizzano la membrana a un livello minimo, il cosiddetto livello di soglia, si innesca il potenziale di azione. Se la depolarizzazione non raggiunge la soglia, non si avvia il potenziale di azione, e il potenziale graduato si estingue. Poiché la depolarizzazione è necessaria per eccitare la cellula e avere un potenziale di azione, un potenziale graduato depolarizzante si dice potenziale postsinaptico eccitatorio (EPSP). Un potenziale graduato iperpolarizzante allontana il potenziale di azione dal valore di soglia (IPSP). La maggiore intensità iniziale di un potenziale graduato eccitatorio, fa sì che esso giunga soprasoglia presso al cresta assonale, quindi viene iniziato un potenziale d’azione.
Se molti stimoli arrivano simultaneamente, i loro potenziale graduati si sommano: diversi potenziale graduati eccitatori sottosoglia possono sommarsi per dare un potenziale soprasoglia. L’avvio di un potenziale d’azione a partire da diversi potenziale graduati sottosoglia simultanei è noto come sommazione spaziale. Non tutte le sommazioni provocano un potenziale di azione: un potenziale graduato iperpolarizzante si oppone ad un potenziale graduatodepolarizzante, dando un segnale sottosoglia.
Due potenziali graduati sottosoglia in rapida successione si sommano se arrivano alla zona trigger abbastanza vicini nel tempo: sommazione temporale.
I potenziali di azione sono segnali elettrici rapidi che corrono lungo l’assone fino al suo terminale. I potenziali di azione sono identici tra loro e non perdono di intensità viaggiando lungo la cellula: il potenziale di azione all’estremità assonale distale è identico al potenziale di azione partito dalla zona trigger. L’intensità del potenziale graduato che avvia un potenziale di azione non incide sul potenziale di azione, purché il potenziale graduato sia soprasoglia. Poiché i potenziali di azione si verificano come depolarizzazioni massimali oppure non si verificano affatto sono definiti anche fenomeni tutto o nulla.
I potenziali d’azione sono variazioni di potenziale di membrana legate all’apertura di canali voltaggio-dipendenti, che modificano la permeabilità della membrana al Na e al K. Si hanno tre fasi: fase ascendente del potenziale di azione, fase discendente, e fase di iperpolarizzazione postuma. Prima e dopo il potenziale di azione il neurone si trova al potenziale di membrana al riposo (-70mV).
La fase ascendente è legata ad un temporaneo aumento della permeabilità cellulare all’Na. Il potenziale di azione comincia quando un potenziale graduato che raggiunge la zona trigger depolarizza la membrana fino al valore soglia. Quando la cellula si depolarizza, i canali per l’Na voltaggio dipendenti si aprono e la membrana diviene bruscamente molto più permeabile all’Na. L’Na entra facilmente sia perché è molto più concentrato nel liquido extracellulare, sia perché è attratto dal potenziale di membrana intracellulare negativo. L’arrivo di cariche positive nel liquido intracellulare depolarizza la cellula, cioè la rende più positiva. Quando il potenziale di membrana diventa positivo, non c’è più carica elettrica che attragga l’Na nella cellula. Rimane tuttavia il gradiente di concentrazione dell’Na, e lo ione continua ad entrare. Al picco della fase ascendente, la membrana ha cambiato polarità: il versante intracellulare è più positivo di quello extracellulare. Questo cambiamento è rappresentato dall’overshoot, cioè la porzione di potenziale di azione che è al di sopra degli 0mV. Gli ioni sodio continuano a diffondere nel neurone, portando il potenziale di membrana verso il valore del potenziale di equilibrio per l’Na (+60mV). La permeabilità al sodio cala rapidamente e il potenziale di azione raggiunge il picco a +30mV. La fase discendente è legata ad un aumento di permeabilità per il potassio: i suoi canali voltaggio dipendenti si aprono in risposta alla depolarizzazione, ma sono più lenti. Quando i canali per il K si aprono il potenziale di membrana ha già raggiunto i +30mV. Quando i canali per l’Na si chiudono, i gradienti elettrico e di concentrazione del K, favoriscono l’uscita di questo ione dalla cellula. Il potenziale di membrana diventa più negativo, provocando la fase discendente e portando la cellula verso il suo potenziale di riposo. I canali del K si chiudono lentamente e, invece di ripolarizzarsi a -70 mV, la cellula si iperpolarizza fino a -90 mV. L’ulteriore perdita di K genera una iperpolarizzazione postuma Quando i canali per il K si chiudono si ripristina una normale permeabilità al K, qualche ione Na rientra nella cellula, riportando il potenziale a -70 mV.
Il potenziale di azione è una variazione del potenziale di membrana che si ha quando si aprono i canali ionici, aumentando la permeabilità cellulare prima all’Na poi al K. L’ingresso di Na depolarizza la cellula. Questa depolarizzazione è seguita da una uscita di K, che riporta la cellula al potenziale di membrana a riposo.
I canali per il Na voltaggio dipendenti hanno due cancelli che regolano il movimento ionico, detti di attivazione e di in inattivazione, che oscillano tra due posizioni, per aprire e chiudere i canali dell’Na.
Quando il neurone si trova al potenziale di riposo, il cancello di attivazione è chiuso e l’Na non entra, mentre il cancello di in inattivazione è aperto. Quando la membrana cellulare si depolarizza, il cancello di attivazione si apre e il canale dell’Na si apre. L’ingresso di cariche positive depolarizza l’interno della cellula e genera un circuito da feedback positivo, quindi sempre più canali per l’Na si aprono. Fino a che la cellula rimane depolarizzata, i cancelli di attivazione sono aperti. Per interrompere la crescente depolarizzazione cellulare c’è bisogno del secondo cancello. Il breve lasso di tempo in cui il cancello è aperto è sufficiente a creare la fase ascendente del potenziale di azione. Dopo che questa è iniziata, il cancello di inattivazione dell’Na si chiude, e diminuisce la permeabilità all’Na , l’ingresso di questo ione si ferma, e il potenziale di azione raggiunge il picco. Quando il neurone si depolarizza durante l’uscita di K, i canali per l’Na si riposizionano in modo da poter rispondere alla successiva depolarizzazione.
Lezione 6 (presa quasi intermente dagli appunti di Marco visto che ha ripetuto e ripetuto e ripetuto…le stesse cose)
L’altra volta abbiamo visto come a livello somatodendritico si può produrre una depolarizzazione nel neurone che si propaga fino alla zona trigger, ricca di canali del sodio voltaggio dipendenti. Se la depolarizzazione è soprasoglia, si ha un’apertura dei canali che creano un potenziale di azione. Abbiamo visto che il flusso dei vari ioni, attraverso la membrana, è determinato oltre che dai canali aperti, anche da due fattori: il gradiente di concentrazione ed il potenziale di membrana. Supponiamo che il potenziale di membrana sia -100mV. Supponiamo che il potassio è all’equilibrio perché le due forze si bilanciano ed annullano. Il potassio, che lungo il suo gradiente di concentrazione si muoverebbe, è in realtà trattenuto dalla barriera del potenziale elettrico.
Se diminuisce il potenziale elettrico, allora la forza del gradiente di concentrazione ha il sopravvento, ed il potassio può fluire. Quando il potenziale di membrana è all’equilibrio, il potassio non si muove. Se il potenziale di membrana varia, e il flusso ionico è consentito dall’apertura dei rispettivi canali, questi flussi si possono verificare. Nel caso del potenziale di azione, inizialmente, prima della depolarizzazione, il potenziale è quello di riposo, pari a -70mV. Se la depolarizzazione arriva alla zona trigger innesca l’apertura dei canali sodio e la membrana si depolarizza. Il sodio entra sia per mezzo del gradiente di concentrazione sia per mezzo del gradiente di potenziale elettrico. Ogni ione, se potesse, eguaglierebbe il valore del potenziale di equilibrio ricavato dall’equazione di Nerst. Il sodio sarebbe portato a fluire all’interno ma solo se abbiamo pure un gradiente di potenziale elettrico che ne permette tale movimento. Lo ione sodio è attratto dalle cariche negative situate all’interno della membrana. Questo è quello che avviene nel potenziale di azione. Se i canali si aprissero e la permeabilità del sodio fosse elevata, avremo una membrana con un potenziale di +65 mV. Tale evento nella fase dell’overshoot in parte si verifica. Quando la permeabilità del sodio aumenta, il potenziale cresce in valore fino a +30mV raggiungendo quasi il potenziale di membrana di equilibrio del sodio ottenuto dall’equazione di Nerst.
Quando arriva la depolarizzazione vengono innescati due fenomeni che però temporalmente procedono in maniera diversa. Abbiamo l’apertura rapida dei canali del sodio e, a mano a mano che la zona trigger si depolarizza, lentamente, ma molto lentamente cominciano ad aprirsi anche i canali del potassio. La permeabilità del potassio aumenta perché si aprono oltre ai normali canali del potassio, anche altre vie di passaggio del potassio come i canali ionici del potassio voltaggio dipendenti. A questo punto il potassio è libero di spostarsi a cavallo della membrana. Se il potenziale di membrana fosse quello di riposo, il potassio sarebbe già all’equilibrio e non si sposterebbe. Se la membrana si è però depolarizzata il potassio non è più all’equilibrio. Il potassio esce quando la membrana si è già depolarizzata perché la cinetica dei canali potassio è più lenta dei canali del sodio. Mentre i canali del sodio si aprono rapidamente i canali del potassio si aprono con una cinetica molto più lenta. Inizialmente il potassio è all’equilibrio ed il sodio può entrare, cosa che fa. Se si aprissero dei canali che lasciassero passare contemporaneamente sodio e potassio in egual quantità, che succederebbe? Il potenziale di membrana arriverebbe intorno a zero se la permeabilità fosse la stessa. Il potenziale di membrana sarebbe all’incirca la somma dei potenziali dei due ioni. Sarebbe intorno a -5mV perché sommeremo -70mV per gli ioni K+ e +65mV per gli ioni Na+. I canali che si aprono a livello della zona somatodendritica sono canali a porta chimica che consentono il passaggio sia di potassio che di sodio. Pur essendo data la possibilità ai due ioni di muoversi nella membrana abbiamo per il sodio una grossa forza che lo trascina all’interno mentre per il potassio avremo una forza più debole che lo trascina all’esterno della cellula. Se aprissimo quindi contemporaneamente un canale che lascia passare sodio e potassio, il potassio si muoverebbe solo lungo gradiente di concentrazione più lentamente rispetto al sodio che fluirebbe molto più rapidamente perché spinto sia dal gradiente di concentrazione che dal potenziale elettrico. La membrana si depolarizzerebbe ed il fenomeno proseguirebbe fin tanto che non si raggiunga una via di mezzo che equilibri la fuoriuscita del potassio e del sodio. Visto che le concentrazioni rimangono sempre le stesse, questa condizione si realizza solo quando il potenziale arriva all’equilibrio tra i valori del potenziale del sodio e del potassio. I recettori nicotinici, che sono recettori dell’acetilcolina e canali ionici a porta chimica, sono presenti nelle sinapsi ma anche a livello neuromuscolare. Essi sono recettori canale che lasciano passare sia sodio che potassio. Il loro utilizzo porta ad una depolarizzazione della membrana dato che il sodio entra esplosivamente nella cellula più del potassio che invece esce dalla cellula. La generazione dei potenziali di azioni deve essere quindi controllata. Se un neurone deve rilasciare molti neurotrasmettitori farà variare la frequenza dei potenziali di azione. Molte tossine e anestetici agiscono sull’apertura dei canali del sodio e del potassio. Molti anestetici si legano ai canali del sodio voltaggio dipendenti ed in questo modo bloccano l’insorgenza del potenziale di azione in quei neuroni che generano la sensazione del dolore. Altri anestetici, come il cloroformio, agiscono sui canali del potassio. Una depolarizzazione della membrana provoca una perdita di funziona del neurone se essa è costante, cosa che dovrebbe avvenire se blocchiamo i canali del potassio. Le depolarizzazioni della membrana sono importanti sia per i tessuti eccitabili che per gli altri tessuti.
Possiamo distinguere adesso tra una cellula eccitabile ed una non eccitabile. Tutte le cellule hanno un potenziale di membrana. Tutte le cellule possono variare il loro potenziale di membrana modulando lo stato di apertura dei loro canali ma, solo i tessuti eccitabili possono produrre un potenziale di azione che si propaga lungo la superficie della cellula. Questa è la differenza tra tessuto eccitabile e non eccitabile. Molte cellule ghiandolari secernono prodotti a seguito della depolarizzazione della membrana. La membrana si depolarizza aprendo i canali del calcio (non necessariamente voltaggio dipendenti) portando alla secrezione del prodotto di quella cellula. Non è quindi necessario che si scateni un potenziale di azione. Queste cellule, anche se non sono eccitabili, compiono alcune delle loro funzioni attraverso la modulazione del potenziale di membrana. Cellule nervose, come i neuroni, il muscolo scheletrico ed il muscolo cardiaco sono i tre principali tipi di tessuti eccitabili.
L’overshoot si ha quando il potenziale di membrana si inverte di polarità a causa del passaggio degli ioni sodio. Se la permeabilità non si attenuasse, il potenziale dovrebbe approssimarsi a quello del sodio ma, dopo un picco a circa 30mV, avremo una ripolarizzazione della membrana a causa dell’apertura dei canali del potassio. Sembra che l’apertura dei canali sia innescata dalla depolarizzazione. I diversi tempi sono dati da una diversa cinetica dei canali del sodio e del potassio. In un millisecondo i canali del sodio si aprono e chiudono. La ripolarizzazione avviene perché i canali del sodio cominciano a chiudersi e perché aumenta “lentamente”, ma progressivamente, la permeabilità del potassio. Quando raggiungiamo il picco comincia la chiusura dei canali del sodio e si aprono invece i canali del potassio che quindi potrà fluire all’esterno. Nel momento in cui il potassio può fluire liberamente, trova una membrana depolarizzata che gli permette di fluire fortemente dall’interno all’esterno della cellula. Al picco del potenziale di azione la polarità è invertita. Il potassio quindi sarà spinto non solo dal suo gradiente di concentrazione ma anche dal gradiente del potenziale elettrico. Il potassio si muoverà fino a ripristinare il suo potenziale di equilibrio. Vi sarà anche una iperpolarizzazione della membrana quando i canali del sodio si saranno chiusi completamente e quando la permeabilità del potassio raggiungerà un valore più elevato di quello che si ha nello stato di riposo. L’eccesso di ioni potassio che escono porterà quindi ad una iperpolarizzazione. Successivamente i canali del potassio cominciano a chiudersi e vengono ristabiliti lentamente i valori del potenziale di membrana a riposo anche grazie all’attività della pompa sodio-potassio ATPasi. Il potenziale di membrana a riposo è dato da pori del potassio che lasciano sempre passare il potassio. Le cellule possono variare temporaneamente il loro potenziale di membrana tramite la modulazione di particolari canali del potassio che non sono i pori. L’apertura e chiusura dei canali determinano la refrattarietà. Un tessuto eccitabile nel quale si sia prodotto un potenziale di azione, non può produrre immediatamente un nuovo potenziale di azione anche se arriva uno stimolo nuovo. Il doppio cancello dei canali per il sodio ha un ruolo importante nel fenomeno noto come periodo refrattario. La refrattarietà di un neurone si riferisce al fatto che, una volta che sia stato avviato un potenziale di azione, un secondo potenziale di azione non può partire, per quanto intenso sia lo stimolo: questo è definito periodo refrattario assoluto. Questa proprietà garantisce che un secondo potenziale di azione non partirà prima che il precedente si sia esaurito. I potenziali di azione non possono sovrapporsi a causa dei periodi refrattari, quindi non possono sommarsi, come fanno i potenziali graduati. Dopo che i canali per l’Na si sono riposizionati, ma prima che la membrana sia nuovamente al potenziale di riposo, un potenziale graduato più intenso del normale può avviare un altro potenziale di azione: periodo refrattario relativo. L’Na che entra è bilanciato dal K che ancora esce: i flussi opposti di carica si equilibrano, e il potenziale di membrana non arriva alla soglia: per questa ragione un potenziale graduato deve essere più intenso del normale per provocare un potenziale di azione. Il periodo refrattario limita la frequenza di trasmissione dei segnali lungo il neurone. Inoltre il periodo refrattario assoluto assicura l’unidirezionalità di un potenziale di azione dal soma al terminale assonale, impedendo che esso venga condotto all’indietro lungo le zone della membrana che ha appena percorso.
In tutto il periodo in cui vige l’iperpolarizzazione, una cellula può produrre un altro potenziale di azione solo se lo stimolo è maggiore di quello che ha iniziato la prima depolarizzazione. Il potenziale di azione è quello di una depolarizzazione che si propaga dalla zona trigger lungo l’assone fino all’estremità sinaptica. Nei pressi del bottone sinaptico, prima che l’assone del neurone prenda contatto con un altro neurone, il potenziale di azione arriva ed innesca l’apertura di canali del calcio voltaggio dipendenti. Questi canali fanno fluire calcio dal mezzo extracellulare all’interno determinando il rilascio per esocitosi di neurotrasmettitori. Questo potenziale, quindi, determina in ultima analisi l’ingresso del calcio e il rilascio di neurotrasmettitori. Da un segnale elettrico si genera pertanto un segnale chimico. Nell’assone gigante di calamaro si ha la stessa composizione di canali del sodio e potassio voltaggio dipendenti della zona trigger. Quello che abbiamo visto accadere nella zona trigger avviene lungo tutto l’assone. Una volta generato un potenziale di azione a livello della zona trigger, la depolarizzazione si propaga a valle dell’assone. Qui la depolarizzazione trova nuovi canali voltaggi dipendenti, che aprendosi continuano la propagazione del potenziale. Il potenziale di azione prodotto al livello della zona sottostante della zona trigger, a sua volta induce una depolarizzazione che si propaga localmente a valle. A valle noi avremo una depolarizzazione crescente e locale della membrana che, arrivando soprasoglia, scatena un potenziale di azione più a valle e così via. In questo modo il potenziale arriva fino all’estremità sinaptica. Tutto questo può avvenire però solo se la “miccia” (zona trigger) si è accesa. In un invertebrato l’assone ha le stesse caratteristiche della zona trigger.
Fonte: http://sommofabio.altervista.org/ANNO2/Fisiologia/Fisiolgia-Mauro04-Sistema_nervoso1.doc
Autore del testo: non indicato nel documento di origine
SISTEMA NERVOSO PERIFERICO
Il sistema nervoso periferico è costituito dall’insieme dei nervi, cordoni di fibre nervose che si distribuiscono a tutto il corpo. La maggior parte dei nervi emerge dal nevrasse (nervi spino-encefalici o cerbro-spinali): viene suddivisa in nervi spinali e nervi cranici o encefalici, a seconda che i detti nervi emergano dal midollo spinale o dall’encefalo. I nervi emergenti dai gangli simpatici, detti nervi simpatici, fanno parte del sistema nervoso simpatico.
Nervi spino-encefalici – sono deputati alla sensibilità generale e specifica ed alla motilità della muscolatura volontaria, in quanto contengono sia fibre sensitive che fibre motrici somatiche. Distinguiamo:
nervi contenenti sole fibre sensitive – nervi sensitivi;
nervi contenenti sole fibre motrici somatiche – nervi motori;
nervi contenenti sia fibre sensitivhe che fibre mostrici somatiche – nervi misti
I nervi motori, destinati ad innervare i muscoli, contengono anche alcune fibre sensitive per le etrminazioni sensitive dei muscoli stessi. I nervi spino-encefalici possono inoltre accogliere, per un certo tratto del loro percorso, fibre pregangliari e fibre postgangliari del simpatico.
Nervi simpatici – sono deputati all’innervazione dei visceri, sia all’attività della muscolatura involontaria che all’attività secretoria ghiandolare; sono costituito maggiormente da fibre post-gangliari ed alcuni da fibre pre-gangliari. Provvedono anche all’innervazione sensitiva dei visceri per mezzo di fibre sensitive che ricevono dai nervi spino-encefalici.
Origine dei nervi
Origine reale – è la sede in cui si trovano i neuroni dai quali derivano le fibre che costituiscono il nervo stesso;
Fibra nervosa |
Origine reale |
Fibre sensitive |
Gangli spino-encefalici o cerebro-spinali costituiti dai neuroni sensitivi a T, che hanno un neurite che si divide in una fibre periferica che termina alla periferia del corpo dove forma terminazioni sensitive ed in una fibra centrale che penetra nel nevrasse |
Fibre motrici somatiche |
Nuclei somatomotori del nevrasse, costituiti dai motoneuroni |
Fibre pre-gangliari |
Nuclei viscero-effettori del nevrasse, costituiti dalle cellule effettrici viscerali |
Fibre post-gangliari |
Gangli simpatici, costituiti dai neuroni gangliari simpatici |
Origine apparente – è la sede in cui il nervo ha apparentemente inizio: per un nervo spino-encefalico è il suo punto di emergenza dal nevrasse, per un nervo simpatico è il ganglio simpatico dal quale il nervo origina.
Al sistema nervoso periferico appartengono inoltre i gangli, aggregati neuronali dislocati esternamente al nevrasse:
gangli spino-encefalici o cerbro-spinali – posti lungo il decorso dei nervi spino-encefalici, rappresentano le origini reali delle fibre sensitive dei nervi stessi;
gangli simpatici – rappresentano le origini reali delle fibre post-gangliari dei nervi simpatici.
fonte: http://digilander.libero.it/fisioterapiad/sistema%20nervoso%20periferico.doc
Autore del testo: non indicato nel documento di origine
Il sistema nervoso si può dividere in periferico e centrale.
Il sistema nervoso centrale nei vertebrati è posto all’interno di strutture ossee che ne assicurano la salvaguardia.
Una parte di esso è posto infatti all’interno del midollo osseo lungo i l quale passano molte terminazioni nervose, mentre l’encefalo, contenuto nel cranio, è formato da diversi tipi di centri nervosi che paragonano ed integrano ed elaborano informazioni provenienti dal sistema nervoso periferico per poi inviare i risultati ai muscoli ed a tutte le parti del corpo tramite il sistema nervoso periferico.
Sistema nervoso periferico
Ne fanno parte i neuroni sensori che hanno il compito di portare le informazioni al sistema nervoso centrale, a differenza dei neuriti motori che uscendo dal sistema nervoso trasmettono gli ordini ai muscoli e alle ghiandole.
I neuriti ed i dendriti dei neuroni motori e sensori sono riuniti in fasci detti nervi se esterni al sistema nervoso e tratti se interni.
I neuroni sensori sono raggruppati in gangli nervosi .I neuroni che trasportano le informazioni dal sistema periferico a quello centrale sono detti neuroni afferenti, al contrario di quelli che compiono il percorso opposto (neuroni efferenti).
Il sistema nervoso periferico si può dividere in somatico ed autonomo o vegetativo.
Il sistema somatico comprende prolungamenti di neuroni sensori e motori che tengono in collegamento il sistema encefalico con muscoli e ghiandole ed è quindi responsabile di movimenti muscolari volontari.
Il sistema vegetativo o autonomo invece raggruppa nervi e tratti che usa indipendentemente dalla volontà essendo infatti controllato da un sistema interno all’encefalo: l’ipotalamo ed agisce ad esempio sulla muscolatura liscia (involontaria) di stomaco, e polmoni.
Il sistema nervoso autonomo comprende inoltre il sistema simpatico e quello parasimpatico che interessano stessi organi con effetti opposti tra loro.
Il sistema simpatico in caso di allarme stimola il cuore ad aumentare i battiti, ed una maggiore immissione di ossigeno .
Mentre il parasimpatico è coinvolto nelle fasi di riposo e rallenta i battiti durante il sonno.
Sistema nervoso centrale
Il midollo spinale contiene le vie di comunicazione nervosa. Sia gli interneuroni sia i neuroni motori costituiscono una pila detta materia grigia e posta sotto il cervello vero e proprio.
Mentre i neuriti degli interneuroni sensori e motori sono coperti da mielina biancastra e sono detti sostanza bianca.
L’encefalo è racchiuso invece all’interno del cranio. È costituito da:
- tronco celebrare che è una continuazione del midollo spinale.
È composto dal bulbo (midollo allungato) ed ha funzioni di coordinare i riflessi motori insieme al cervelletto col quale è collegato da peduncoli cerebellari.
Le fibre sensorie che passano all’interno si incrociano provocando la risposta alla parte destra del corpo dalla parte sinistra del cervello e viceversa.
Nel tronco ci sono i centri riflessi che regolano attività involontarie ed i centri dell’automatismo che regolano attività quali i movimenti respiratori ed il ritmo cardiaco.
- il cervelletto ha forma simile al cervello. La parte inferiore elabora segnali provenienti dal labirinto(la parte interna dell’orecchio responsabile dell’equilibri) inviando tutti i dati relativi alla posizione del corpo al cervello.
Mentre la parte superiore coordina i movimenti di grande precisione.
- sistema reticolare
lungo il tronco celebrale l’intreccio di tanti neuroni forma il sistema reticolare collegato al talamo. Il sistema reticolare fa da filtro degli stimoli sensori, identificando e dando la precedenza ai più importanti.
- Talamo
Il talamo è posto sopra il tronco celebrale e ha compito di suddividere le informazioni in arrivo dai recettori sensori, è inoltre il centro di smistamento delle sensazioni di dolore ed è in parte responsabile delle emozioni.
- Ipotalamo (produce ormoni)
Si trova sotto il talamo e controlla l’omeostasi (mantenimento equilibrio interno al corpo), controlla le funzioni involontarie fondamentali come fame e sete.
Per quanto riguarda la sete c’è lo stimolo di bere nel momento in cui i ricettori rilevano una minore presenza di Sali nel sangue. Invece arriva lo stimolo della face quando il centro di sazietà e appetito stimolano o meno a mangiare a seconda dello zucchero presente nel sangue.
- sistema libico
Rete di neuroni estesa sotto il cervello che comprende encefalo talamo ed ipotalamo.
Qui hanno origine tutti quei comportamenti derivati da sensazioni visive o uditive come paura e gioia.
- CERVELLO
La parte superiore del cervello presenta ripiegamenti per poter aumentare il numero di neuroni e ridurre lo spazio). Esso è costituito da una parte superficiale detta corteccia celebrale (composta da sostanza bianca e materia grigia), essa è composta da un gran numero di neuroni responsabili dei movimenti volontari. Il cervello è diviso in due emisferi, inoltre sono presenti molti solchi che lo dividono in vari Lobi, il più importanti di questi solchi è il solco di Rolando, nella zona anteriore a questo solco sono posti tutti i centri che coordinano i movimenti volontari: l’insieme di questi cerchi formano la corteccia motoria.
Invece nella zona posteriore al solco ci sono i centri di elaborazioni di stimoli sensori provenienti dalle varie parti del corpo, l’insieme di questi centri forma la Corteccia Sensoria Associativa, caratterizzata dagli interneuroni che integrano gli stimoli sensori con la memoria e le emozioni ed organizzano le idee.
Emisferi celebrali
L’emisfero destro comanda la parte sinistra del corpo perché i tratti nervosi si incrociano nel tronco celebrale. Le due parti appaiono identiche e collegate da una struttura detta corpo calloso. L’emisfero sinistra controlla invece il linguaggio, la capacità di leggere e scrivere ed eseguire calcoli. Mentre la perdita delle funzione dell’emisfero destro hanno causato l’impossibilità di vedere a tre dimensioni, la perdita di talento musicale, l’impossibilità di riconoscere forme e di risolvere problemi.
Meccanismi di collegamento
Alla base del funzionamento del sistema nervoso c’è la capacità di una singola cellula di avvertire cambiamenti nell’ambiente e rispondere nel modo adatto allo stimolo. Ad esempio le alghe unicellulari seguono la luce se la percepiscono, a differenza dei vermi piatti nei quali la presenza di neuroni ed interneuroni permette di scegliere tra diverse possibilità di comportamento. Gli anellidi. Hanno invece già formato l’enecefalo frontale e catene di gangli nervsi.
L’evoluzione non fù rilevante né per il sistema nervoso centrale né per il tronco celebrale, mentre cervelletto, tetto ottico, ed encefalo hanno subito variazioni: il cervelletto ad esempio si è evoluto per il controllo di movimenti più precisi solo in uccelli e mammiferi. Mentre il tetto ottico che elabora impulsi ottici si è ampliato in pesci anfibi e uccelli. Il proencefalo (stimoli olfattivi) si è evoluto solo in parte nei rettili. Per quanto riguarda la corteccia celebrale presente nei mammiferi è inesistente in pesci e anfibi e abbozzato in rettili e uccelli.
Fonte: http://www.sunfire.altervista.org/Materie/Julia/Biologia/Teoria/Sistema%20nervoso%20centrale%20e%20periferico.doc
Autore del testo: non indicato nel documento di origine
Il sistema nervoso autonomo o vegetativo è così chiamato perché rappresenta quella parte del sistema nervoso che, godendo di una certa autonomia nei confronti del nevrasse, esercita la sua attività di controllo sulle funzioni vegetative in modo automatico ed inconsapevole.
Esso è diviso in due grandi strutture: il simpatico (o ortosimpatico) ed il parasimpatico (Cattaneo L., 1989). La sostanziale differenza tra i due sistemi è soprattutto di ordine funzionale in quanto, pur essendo ambedue presenti nei diversi organi, svolgono un’azione antagonista.
Il sistema simpatico attiva la risposta dell'organismo ad una situazione di allarme, lotta e stress. Esso, quando viene attivato, produce una serie di effetti quali: accelerazione del battito cardiaco, dilatazione dei bronchi, aumento della pressione arteriosa, vasocostrizione periferica, dilatazione pupillare, aumento della sudorazione. I mediatori chimici di queste risposte vegetative sono la noradrenalina, l'adrenalina, la corticotropina e diversi corticosteroidi.
Al contrario, il sistema nervoso parasimpatico, quando viene attivato, produce: rallentamento del ritmo cardiaco, aumento del tono muscolare bronchiale, dilatazione dei vasi sanguinei, diminuzione della pressione, rallentamento della respirazione, aumento del rilassamento muscolare, respiro più calmo e profondo, genitali, mani e piedi più caldi. Esso agisce attraverso il mediatore chimico acetilcolina. Il sistema parasimpatico rappresenta la normale risposta dell'organismo ad una situazione di calma, riposo, tranquillità ed assenza di pericoli o stress.
http://web.tiscali.it/codialogos/SNA%20E%20DISAGIO%20PSICHICO.doc
Autore del testo: non indicato nel documento di origine
Sistema nervoso centrale autonomo e periferico
Visita la nostra pagina principale
Sistema nervoso centrale autonomo e periferico
Termini d' uso e privacy