Tessuto nervoso
Tessuto nervoso
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Le informazioni di medicina e salute contenute nel sito sono di natura generale ed a scopo puramente divulgativo e per questo motivo non possono sostituire in alcun caso il consiglio di un medico (ovvero un soggetto abilitato legalmente alla professione)
Tessuto nervoso
generalità
Il tessuto nervoso è composto da:
cellule nervose o neuroni, unità morfologiche e funzionali del sistema nervoso
cellule della nevroglia: svolgono il ruolo che negli altri organi è svolto dal tessuto connettivo, non presente nel sistema nervoso
a scopo didattico descrittivo in una cellula nervosa si possono distinguere alcuni distretti
le parti in cui si può distingure il neurone sono:
il corpo cellulare o pirenoforo o soma: centro metabolico del neurone caratterizzato dalla presenza del nucleo e degli organuli citoplasmatici
i dendriti: arborizzazioni in genere molto numerose originate dal corpo cellulare come vere e proprie espansioni della cellula;
costituiscono l’apparato di ricezione del neurone.
Da un punto di vista funzionale conducono l’impulso nervoso verso il corpo cellulare (conduzione cellulipeta o centripeta).
assone: unico, appare di solito come prolungamento nettamente definito.
Da un punto di vista funzionale conduce l’impulso nervoso lontano dal corpo cellulare (conduzione cellulifuga o centrifuga).
le cellule nervose sono in rapporto tra di loro e formano reti estremamente complicate: il rapporto che si viene a stabilire tra parti di cellule è un rapporto di contiguità (non di continuità morfologica) definito sinapsi
Proprietà della cellula nervosa
le cellule nervose sono accomunate dalle seguenti proprietà:
Irritabilità: capacità di stabilire una d.d.p. tra interno ed esterno, (da -70 a + 20,+30 mV).
Conducibilità: proprietà di propagare l'impulso lungo tutta la membrana.
comunicazione: l’impulso può essere trasferito mediante sinapsi (chimiche) o nexus (elettriche) ad altre cellule.
secrezione: (neurotrasmettitori, neuromodulatori etc.)
Perdita della capacità replicativa (cellula perenne?)
Classificazione delle cellule nervose
Si è cercato di raggruppare le cellule nervose in base a dei criteri comuni ad altri tessuti
Esiste difficoltà dovuta al fatto che le cellule nervose possono avere forma e dimensioni estremamente variabili.
il criterio utilizzato tiene conto sia di aspetti morfologici che funzionali.
Si distinguono 3 categorie di cellule nervose:
sensitive
neuroni bipolari
neuroni multipolari
Le cellule sensitive sono i cosiddetti recettori di senso specifico; si trovano negli organi di senso.
Sono cellule altamente differenziate, predisposte per percepire stimoli di varia natura, (luminosi, onde sonore, segnali chimici ecc.).
Le cellule sensitive primarie (olfattive e le visive) sono dotate di un prolungamento che corrisponde all’assone.
Le cellule sensitive secondarie (acustiche, statocinetiche (dell’equilibrio) e gustative) non hanno un proprio prolungamento ma si mettono in comunicazione con prolungamenti provenienti da altre cellule nervose stabilendo con esse collegamenti sinaptici che vengono definiti giunzioni citoneurali o neurosensoriali.
Le cellule sensitive primarie, avendo un prolungamento nervoso proprio, sono da considerare neuroni a tutti gli effetti, neuroni unipolari.
Le cellule sensitive secondarie, che non hanno un proprio prolungamento, appartengono ai cosiddetti epiteli sensoriali.
Neuroni Bipolari
I neuroni bipolari costituiscono i neuroni afferenti che si trovano nei gangli spinali e nei gangli o nei nuclei dei nervi cranici.
Questi neuroni si dicono bipolari in quanto sono dotati di due prolungamenti, identici dal punto di vista morfologico, con i caratteri tipici di una assone.
Dal punto di vista funzionale uno dei due prolungamenti conduce l’ impulso in direzione centripeta e uno in direzione centrifuga.
durante la vita embrionale i neuroni bipolari hanno i due prolungamenti in posizione opposta e prendono il nome di neuroni bipolari oppositopolari
nell’organismo adulto hanno i prolungamenti dalla stessa parte rispetto al corpo cellulare e per un certo tratto essi decorrono appaiati per poi allontanarsi in uguale direzione ma con verso opposto, sono i cosiddetti neuroni bipolari pseudounipolari a T.
esiste anche una forma intermedia tra la forma embrionale e la forma adulta: viene detta neurone pseudounipolare a V nella parte iniziale i due prolungamenti non sono completamente appaiati.
L’ avvicinamento dei due prolungamenti ha un preciso significato funzionale: il segnale passa direttamente dal prolungamento dendritico a quello assonico senza dover percorre tutto il corpo cellulare.
Nell’organismo adulto permangono neuroni bipolari oppositopolari a livello dei gangli annessi all’ ottavo paio di nervi encefalici. (l’avvicinamento dei due prolungamenti non avviene perchè i neuroni sono contenuti in una matrice che formerà dell’ osso, e ciò impedisce l'avvicinamento dei due prolungamenti.
Neuroni multipolari
I neuroni mutlipolari presentano numerosi prolungamenti (fino a varie decine di migliaia) tutti quanti rappresentati da dendriti eccetto uno che è l’assone.
Si distinguono i neuroni multipolari
ad assone lungo o del I tipo di Golgi
ad assone breve o del II tipo di Golgi
Nella classificazione non si tiene conto dell’effettiva lunghezza dell’assone bensì dei rapporti che esso ha con le varie parti del sistema nervoso:
1° tipo del Golgi: gli assoni escono dalla sostanza grigia
2° tipo del Golgi: l'assone non esce mai dalla sostanza grigia.
Morfologia del corpo cellulare
A scopo didattico il neurone può essere scomposto in:
corpo cellulare o pirenoforo o soma o cellula nervosa in senso stretto
prolungamenti nervosi.
A livello del corpo cellulare si trovano:
nucleo e organuli tipici di tutte le cellule.
Inclusioni
Pigmento melanico (sostanza nigra e locus coeruleus)
lipofuscina (pigmento ad usura) accumulo di natura lipidica che aumenta con l’avanzamento dell’ età.
Nucleo
Il nucleo è piuttosto grande, voluminoso, di forma rotondeggiante o ellissoidale
caratteristica principale è la scarsa colorabilità, appare chiaro, di aspetto vacuolizzato e al suo interno appare molto evidente la presenza del nucleolo.
Al contrario di quanto ritenuto in passato tale aspetto del nucleo è da correlare ad intensa trasrizione genica
Nonostante la cellula nervosa differenziata sia incapace di riprodursi (?) (sono repressi i geni che codificano per la duplicazione) essa può compiere le funzioni di sintesi.
All’ interno del citoplasma si trovano
Mitocondri: svolgono come sempre funzioni respiratorie.
l’unica differenza da rilevare è che i mitocondri delle cellule nervose, più spesso di quelli delle altre presentano le creste mitocondriali dirette in senso parallelo rispetto all’asse maggiore anziché in senso perpendicolare.
Lisosomi: contengono enzimi proteolitici e idrolitici; non sono abbondanti.
centrioli, a volte una sola coppia, a volte anche due.
si riteneva che i centrioli fossero unicamente responsabili dell’organizzazione delle fibre del fuso mitotico (non necessario nel neurone diffeenziato)
il significato della loro presenza è legato ai centri di organizzazione microtubulare (vedi microtubuli)).
apparato del Golgi: è un sistema di vescicole citoplasmatiche che si trova in vicinanza del nucleo, nelle cellule nervose circonda tutto quanto il nucleo.
nelle cellule nervose ritroviamo anche
tubuli e vescicole del reticolo endoplasmatico liscio (REL)
reticolo rugoso (RER)
I corpi di Nissl
I corpi di Nissl sono presenti nel corpo cellulare e anche nei dendriti, mancano nell’assone (possono essere presenti nella parte iniziale dell’assone o cono di emergenza).
la microscopia elettronica dimostra che i corpi di Nissl sono ribosomi.
I ribosomi possono trovarsi in forma isolata oppure aggregarsi tra loro in forma di poliribosomi oppure distribuirsi sulla superficie del RER.
sono impegnati nella sintesi di proteine strutturali e funzionali:
Strutturali: possono essere rilasciate liberamente nel citoplasma
Funzionali: sono sintetizzate nel RER cosicchè una volta sintetizzate rimangono racchiuse nelle membrane del RER, nel Golgi e poi nelle vescicole; sono compartimentalizzate rispetto al materiale citoplasmatico.
Le proteine strutturali sono quelle che costituiscono il citoscheletro o strutture di membrana (integrine ecc) .
Le proteine funzionali sono proteine enzimatiche impiegate nella sintesi o degradazione di altre sostanze come i mediatori chimici
flusso assonico
lungo l’assone e alla sua terminazione non ci sono corpi di Nissl quindi a tale livello non avviene sintesi proteica.
l’assone necessita di proteine strutturali e proteine funzionali anche alla sua terminazione
Esiste un sistema di trasporto
Il processo di trasferimento prende il nome di flusso assonico e per mezzo di esso un quantitativo di citoplasma pari a tre volte il contenuto del corpo cellulare si trasferisce in un giorno dal corpo cellulare alla terminazione nervosa.
Oggi si conoscono due componenti del flusso assonico:
anterogrado, dal corpo cellulare alla terminazione.
retrogrado, dalla terminazione al corpo cellulare.
Nell’ ambito del flusso anterogrado possiamo distinguere:
Il flusso lento: interessa principalmente le proteine strutturali e si sposta ad una velocità compresa tra 0,3 e 8 mm al giorno ( interviene nel ricambio dei costituenti strutturali e nei processi riparativi dell'assone a seguito di lesioni).
Il flusso veloce: procede alla velocità di 50 400 mm al giorno e interessa le proteine funzionali (enzimi, mediatori chimici ecc. perciò interviene nelle attività funzionali della cellula nervosa).
La componente retrograda procede ad una velocità di 200 400 mm al giorno; riporta al corpo cellulare cataboliti che possono essere riutilizzati dalla cellula.
meccanismo del flusso assonico
La spinta che fa procedere il flusso è dovuta a proteine che scorrono sui microtubuli del citoscheletro
Quello veloce si verifica grazie all’ attività di proteine che funzionano da veri e propri motori citoplasmatici,
la chinesina per il flusso anterogrado
la dineina per il flusso retrogrado.
Questi motori da un lato legano le vescicole contenenti altre proteine e materiali e dall’ altro scorrono su dei veri e propri binari costituiti da microtubuli e dai filamenti intermedi del tessuto nervoso, i neurofilamenti.
I prolungamenti: I dendriti
sono vere e proprie espansioni del corpo cellulare
rappresentano l’apparato ricevente del neurone e sono sede dei contatti sinaptici
la loro funzione è quella di ampliare la superficie della cellula così da rendere possibile un numero più elevato di sinapsi da parte di terminazioni provenienti da altre cellule
originano con un calibro piuttosto ampio poi tendono a degradare diventando molto sottili all’estremità.
sulla superficie dei dendriti ritroviamo escrescenze dette spine o gemme o gemmule, a volte sono l’unico carattere distintivo tra il dendrite e l’assone
presentano ramificazioni secondarie che si dipartono ad angolo acuto
rappresentano i punti dove avviene il contatto sinaptico
a volte un’unica cellula può stabilire decine di migliaia di contatti grazie all’estrema ramificazione.
sulla superficie esterna dei dendriti non sono presenti delle vere e proprie guaine di rivestimento, comunque all’esterno dei dendriti troviamo sempre della nevroglia (cosa che vale anche per il corpo cellulare).
all’interno dei dendriti si ritrovano i corpi di Nissl e sono presenti le neurofibrille disposte in maniera casuale.
I prolungamenti: l’assone
è detto anche cilindrasse o neurite
è unico e presenta poche ramificazioni lungo il suo decorso che formano angoli di 90° con il tronco principale
il calibro dell’assone è uniforme tranne che nella sua parte iniziale, il cono di emergenza, che ha una forma a imbuto.
Il tratto successivo prosegue con un diametro pressoché costante.
la membrana plasmatica dell’assone prende il nome di assolemma.
la superficie dell’assone è liscia, non presenta le spine e al suo esterno troviamo delle guaine con il compito di isolare l’assone e di favorire la progressione del potenziale d’azione.
all’interno dell’assone non sono presenti i corpi di Nissl ad eccezione del cono d’emergenza in particolari situazioni (lesioni assonali)
sono invece presenti neurofilamenti e neurotubuli con disposizione regolare e parallela all'asse maggiore dell’assone.
La fibra nervosa
fibra nervosa: è l’assone rivestito o meno da guaine di due tipi:
la guaina mielinica
la guaina di Schwann o nevrilemma.
a seconda della presenza e della combinazione delle due guaine le fibre nervose possono essere di 4 tipi diversi e questo consente di fare una classificazione.
possiamo distinguere:
assone nudo
fibra completa
fibra mielinica
fibra amielinica
l’assone nudo è l’assone privo di guaine
si trova nella sostanza grigia del sistema nervoso centrale
questa definizione non va presa alla lettera: il concetto di "nudo" vuol dire che non ci sono né guaina mielinica né guaina di Schwann ma un rivestimento di nevroglia (astrociti) è sempre presente in quanto se due assoni entrassero in contatto tra di loro si avrebbe un corto circuito.
la fibra completa ha entrambe le guaine
si trova nel sistema nervoso periferico:
si tratta del tipo di fibra che costituisce in prevalenza i nervi spinali e i nervi cranici.
la fibra mielinica ha solo la guaina mielinica
costituisce la sostanza bianca del sistema nervoso centrale.
la fibra amielinica ha solo la guaina di Schwann
si ritrova in alcuni fasci nervosi che costituiscono le diramazioni del sistema nervoso autonomo
Istogenesi dei vari tipi di fibra nervosa
La fibra completa
durante lo sviluppo embrionale, quando il prolungamento di una cellula di moto, esce dal midollo spinale o quando i prolungamenti dei neuroni afferenti gangliari fuoriescono dai gangli, vengono avvicinati da alcune cellule che derivano dalle creste gangliari definite lemnoblasti
i lemnoblasti rappresentano i precursori della cellula di Schwann, responsabile della formazione di entrambe le guaine
man a mano che i prolungamenti nervosi si allungano vengono avvicinati da nuovi lemnoblasti e ciascun lemnoblasta accoglie l’assone che sta crescendo in una specie di insenatura
a poco a poco il lemnoblasta circonda tutto quanto l’assone e alla fine la membrana che riveste il lemnoblasta si trova raddoppiata laddove i due lembi del lemnoblasta si avvicinano tra di loro al di sopra dell’assone
dove i lembi si uniscono si forma il mesassone con membrana cellulare raddoppiata.
a livello del mesassone comincia una proliferazione della membrana
questa membrana si allunga cosìcchè un lembo si insinua tra il mesassone e il lemnoblasta.
questo processo di avvolgimento si chiama mielinizzazione.
una volta raggiunto il numero di spire desiderato termina l’allungamento del mesassone e il citoplasma all’interno delle spire viene spremuto e sospinto nella parte periferica del lemnoblasta
a questo punto il lemnoblasta è diventato cellula di Schwann.
si sono costituite:
la guaina mielinica, più interna formata dall’accollamento delle membrane delle varie spire
la guaina di Schwann, formata dalla parte periferica della cellula di Schwann che contiene il nucleo e il citoplasma che è stato spremuto a livello della guaina mielinica.
man a mano che l’assone si allunga nuovi lemnoblasti si dispongono in successione al primo e compiono a loro volta il processo di mielinizzazione.
quando l’assone raggiunge la sua lunghezza definitiva è ricoperto per tutta l’estensione dalle due guaine.
La fibra mielinica
in questo caso la guaina mielinica viene formata da un tipo di cellule che appartengono alla nevroglia e si chiamano oligodendrociti
si comportano in maniera diversa rispetto al lemnoblasta:
l’oligodendrocita presenta un corpo cellulare dal quale si originano alcuni prolungamenti ciascuno dei quali si porta in vicinanza dell’assone.
ciascuno dei prolungamenti forma un mesassone che prolifera, formando delle spire;
successivamente il citoplasma viene spremuto dalle spire e anzichè rimanere all’esterno e costituire la guaina di Schwann o nevrilemma torna nel corpo cellulare che è rimasto estraneo al processo di avvolgimento.
Le fibre amieliniche
Nella loro formazione entrano ancora in gioco i lemnoblasti:
circondano gli assoni, anche più di un assone per ciascuna cellula
costituiscono un mesassone per ogni assone ma il mesoassone non prolifera
non ha luogo quindi la mielinizzazione per cui ciascun assone rimane rivestito da un unico strato della membrana del lemnoblasta
questi assoni e le fibre che ne derivano sono più sottili degli altri
Il nervo
un nervo risulta costituito dall’insieme di molte fibre nervose complete (sistema della vita di relazione) e amieliniche (sistema vegetativo).
nella costituzione di un nervo le fibre che lo compongono sono tenute assieme da tessuto connettivo;
all’esterno di un nervo si osserva una capsula di rivestimento formata da un tessuto connettivo fibrillare denso denominata epinevrio.
dall’epinevrio si dipartono dei setti diretti verso l’interno del nervo scomponendolo in tanti fascetti di fibre nervose; tali setti sono costituiti da un connettivo fibrillare sempre più lasso e nel loro insieme costituiscono il perinevrio.
infine dal perinevrio origina una rete delicata di connettivo reticolare che va a rivestire esternamente ogni singola fibra nervosa; questo connettivo più delicato che avvolge ogni singola fibra è detto endonevrio.
le fibre che compongono il nervo possono essere fibre di moto o efferenti e fibre di senso o afferenti (arco riflesso)
i nervi spinali sono in genere nervi misti motori e sensitivi mentre nel caso dei nervi cranici si verifica che alcuni sono quasi esclusivamente motori come il nervo faciale mentre altri sono quasi esclusivamente sensitivi come ad esempio il nervo trigemino
Tecniche per visualizzare il nervo e osservazione al microscopio.
la metodica più frequentemente usata è la colorazione con acido osmico, un fissativo largamente impiegato per la microscopia elettronica e dotato di affinità per i materiali lipidici della componente mielinica colorandoli in nero (le singole fibre danno luogo a tanti anellini scuri)
La colorazione può essere più o meno intensa a seconda dello spessore della guaina ed anche a seconda del livello al quale la fibra viene colpita dalla sezione.
tecnica delle fibre dissociate
si possono studiare fibre dissociate da un nervo (non una vera e propria sezione longitudinale) colorate con acido osmico,
si osservano delle formazioni allungate che corrispondono alle fibre, ciascuna delle quali presenta una porzione centrale colorata meno intensamente e delle linee più scure che la delimitano lateralmente
i nodi di Ranvier
lungo ciascuna fibra, a intervalli regolari, si osservano dei restringimenti a livello dei quali si interrompe la colorazione nera.
questi restringimenti sono detti nodi di Ranvier e corrispondono alla zona dove termina una cellula di Schwann e ne inizia un'altra
una volta individuati i nodi, si vede che in ogni fibra la distanza tra di essi è pressoché costante, questa distanza si chiama internodo o segmento internodale
in media può avere una lunghezza che si aggira intorno ai 100-200 micron
tale lunghezza ha rilievo per quanto concerne gli aspetti fisiologici della fibra (conduzione saltatoria dell’impulso nervoso)
la velocità di conduzione è direttamente proporzionale alla distanza internodale ed anche dal calibro della fibra
colorazione delle fibre nervose con il nitrato di Argento
colorando la fibra con il nitrato d’argento si mettono in evidenza delle formazioni che assumono l’aspetto di croce e sono situate in prossimità di ciascun nodo di Ranvier (croci di Ranvier)
I nodi di Ranvier sono zone ad elevata permeabilità
Questo fatto fu interpretato come un ulteriore prova della teoria saltatoria nella conduzione dell’impulso poichè dimostra che solo nei nodi di Ranvier esiste una permeabilità e quindi solo a questo livello sono consentiti gli scambi ionici necessari per la generazione del potenziale d’ azione
Le strie di Schmidt Lantermann
al microscopio ottico si possono osservare nell’ambito della guaina mielinica colorata con acido osmico delle linee ad andamento obliquo (strie o incisure o fessure di Schmidt-Lantermann)
rappresentano delle zone a livello delle quali il citoplasma non è stato completamente espulso dalle lamelle durante il processo di mielinizzazione
si tratta di canalicoli citoplasmatici che si portano in maniera elicoidale dalle lamelle più esterne verso le più interne e nel complesso somigliano ad una specie di avvolgimento elicoidale che assume una forma ad imbuto
Il significato delle strie è mantenere una comunicazione tra il citoplasma più esterno e uno strato di citoplasma che rimane in prossimità dell’assone (lamella più interna nella quale la mielinizzazione non provoca accollamento delle membrane (strato adassonale)
Le strie consentono il passaggio di sostanze nutritizie dall’esterno fino allo strato adassonale e da qui verso l’assone
la fibra nervosa completa al Microscopio Elettronico a trasmissione
al microscopio elettronico una fibra completa mostra in sezione:
l'assone
procedendo dall’assone verso l’esterno troviamo lo strato adassonale, cioè la lamella più interna da cui non è stato espulso il citoplasma
ancora più esternamente si trovano diverse lamelle formate da strati di membrane accollate (guaina mielinica)
infine lo starto di citoplasma della cellula di Schwann che forma la guaina di Schwann o nevrilemma
aspetto della guaina mielinica al microscopio elettronico a trasmissione
all’ interno della guaina mielinica si trovano linee chiare e linee scure che si alternano tra di loro con regolarità
la linea scura è detta linea densa maggiore (diametro di 30 A°)
la linea o banda più chiara ha uno spessore di 110-130 A°
all'interno della banda più chiara si trova un’ altra linea scura, più sottile, che viene detta linea intraperiodo o anche linea densa minore, ed ha uno spessore di 10 A°
composizione della guaina mielinica
le linee chiare corrispondono ai materiali lipidici accollati tra di loro
i lipidi hanno preso il sopravvento nella guaina nei riguardi delle componenti proteiche di membrana
ciò è necessario poiché la funzione della guaina è quella di permettere il funzionamento dell’assone isolando gli assoni vicini tra di loro: i lipidi sono i più idonei per garantire l'isolamento
le linee scure derivano dall’accollamento degli strati scuri che normalmente si vedono allo studio con il microscopio elettronico sul versante esterno e su quello citoplasmatico della membrana cellulare:
la linea densa maggiore risulterebbe dalla somma degli strati scuri interni e dall’ unione delle lamelle avvicinatesi dopo l’espulsione del citoplasma
la linea densa minore o intraperiodo è il risultato dell’accollamento degli strati esterni
COMPOSIZIONE CHIMICA DELLA MIELINA
La mielina ha una composizione chimica simile a quella delle membrane plasmatiche di altri tipi cellulari e risulta costituita da lipidi e proteine; si differenzia però per qualità e quantità delle due categorie di composti chimici.
I lipidi costituiscono circa il 70% del peso secco della mielina.
Tra i lipidi sono presenti colesterolo, fosfolipidi e glicolipidi.
Tra i glicolipidi si trova un cerebroside che rappresenta il 20% del peso secco totale della mielina.
L'alto contenuto in lipidi rappresenta una barriera per l'acqua e gli ioni idrosolubili quali il sodio e il potassio, favorendo in tal modo la funzione di isolamento della mielina.
Il contenuto in proteine è inferiore a quello di altre membrane cellulari ed, inoltre, esiste un numero minore di classi proteiche.
Sono del tutto assenti le proteine che facilitano il passaggio degli ioni attraverso lo strato bimolecolare lipidico.
Anche questo fatto contribuisce a rendere impermeabile agli ioni la guaina mielinica.
La Sinapsi
Il compito della sinapsi è quello di trasmettere l'impulso da una cellula ad un’altra, rappresenta quindi il punto in cui la cellula stabilisce rapporti di contiguità con un’altra struttura cellulare che può essere:
una nuova cellula nervosa e in tal caso siamo di fronte ad una cosiddetta sinapsi centrale
cellula di altri tessuti (es. muscolare), siamo di fronte ad una sinapsi periferica.
si distinguono le sinapsi anche in:
chimiche: funzionano con l’ impiego di mediatori, sostanze chimiche,
elettriche: sono dei nexus o giunzioni comunicanti o gap junction.
Le sinapsi chimiche centrali
quando si studia la sinapsi da un punto di vista morfologico si deve distinguere:
una struttura pre-sinaptica, a livello della quale si è generato il potenziale d’ azione
il vallo o spazio sinaptico (o fessura sinaptica)
la struttura post-sinaptica, che deve ricevere il segnale
tipi classici di sinapsi a seconda della struttura postsinaptica
la massima parte delle sinapsi centrali presenta una struttura presinaptica rappresentata dal terminale assonico.
A questo livello l’assone subisce diverse ramificazioni, ciascuna delle quali termina con uno slargamento che prende il nome di bottone sinaptico
a seconda del punto in cui il bottone sinaptico prende rapporto con la struttura post-sinaptica si può fare una distinzione delle sinapsi
si dice sinapsi asso-somatica la sinapsi in cui la struttura post-sinaptica è rappresentata dal corpo cellulare di un altra cellula nervosa.
si chiama asso-dendritica la sinapsi nella quale la struttura post-sinaptica è costituita da un dendrite.
si chiama asso-assonica quando il rapporto sinaptico viene costituito tra il terminale di un assone e un altro assone.
ultrastruttura della sinapsi chimica
bottone sinaptico
all’ interno della membrana pre-sinaptica, nel bottone sinaptico, si riscontrano dei mitocondri, neurofilamenti e microfilamenti ma in particolare abbondano le vescicole sinaptiche:
le vescicole sinaptiche hanno un diametro di 500-600 A, sono delimitate da una membrana simile a quella plasmatica e contengono al loro interno uno o più mediatori chimici
Questi ultimi sono costituiti da molecole di varia natura ; tra queste le più studiate sono: adrenalina, acetilcolina, acido gamma-amminobutirrico (GABA) serotonina, endorfine, ossido nitrico ecc.
b) spazio o fessura o vallo sinaptico
è compreso tra la membrana presinaptica e la struttura post-sinaptica (ampiezza 200 a 400 A°)
contiene materiale elttrondenso, simile al glicocalice in genere costituito da glicoproteine
Questo materiale contiene enzimi capaci di degradare in maniera specifica il mediatore chimico.
c) struttura post-sinaptica
a livello della membrana postsinaptica si trovano concentrati dei recettori per i mediatori chimici
tali recettori sono costituiti generalmente da proteine transmembranali spesso disposte a formare dei canali ionici i quali si aprono al legame con il mediatore chimico.
all’interno della membrana post-sinaptica si può trovare un maggiore addensamento di mitocondri e di acido ribonucleico mentre non sono presenti vescicole sinaptiche
cenni sul meccanismo della trasmissione dell'impulso nervoso
Quando giunge il potenziale d’ azione in prossimità della terminazione assonica viene attuato un blocco delle pompe del calcio che in condizioni di riposo tendono ad estromettere calcio dal bottone sinaptico e a concentrarlo maggiormente all’ esterno dell’ assolemma
A seguito di questo blocco gli ioni calcio diffondono all’interno del bottone terminale e il loro ingresso favorisce l’avvicinamento delle vescicole alla membrana pre-sinaptica e l’esocitosi del mediatore chimico nello spazio sinaptico
il mediatore raggiunge la membrana post-sinaptica, si lega al recettore specifico e lo attiva: il canale ionico si apre e aumenta la conduttanza del Na+ ; gli ioni Na+ entrano nella struttura post-sinaptica che inizia a depolarizzarsi: la struttura post sinaptica diventa sede di un potenziale d’azione
segue l’apertura dei canali per il K+ con ripolarizzazione lenta della membrana plasmatica e ripristino del gradiente elettrico transmembrana
la pompa Na/K temporaneamente bloccata riprenderà la sua funzione consentendo di ripristinare anche il gradiente chimico transmembrana
Il potenziale d’azione si trasmetterà lungo la membrana cellulare
ritorno alle condizioni di riposo
per riportare la sinapsi alle condizioni di riposo intervengono diversi meccanismi.
in primo luogo tornano ad essere attive le pompe del calcio che così estromettono gli ioni dal bottone terminale
a livello dello spazio sinaptico entrano in gioco gli enzimi che possono degradare il mediatore chimico, lo scindono nelle sue componenti e lo rendono inattivo
alcune componenti molecolari vengono recuperate all’ interno del bottone
oltre a ciò avviene un recupero di membrana mediante un meccanismo di endocitosi; piccole porzioni di membrana sinaptica si invaginano e formano delle piccole insenature che poi si richiudono di nuovo a formare vescicole.
queste vescicole vengono successivamente riempite con nuovi mediatori mediante processi metabolici di sintesi che possono avvenire anche localmente nel terminale assonico
La nevroglia
la nevroglia rappresenta un tessuto a sé stante che si frappone agli elementi nervosi
riempie gli spazi che si vengono a formare tra le cellule nervose e le loro varie componenti, dendriti e assoni.
nell’ambito del tessuto nervoso svolge le stesse funzioni che il connettivo svolge nei confronti di tutti gli altri tessuti
le funzioni si possono riassumere come:
sostegno meccanico
sostegno trofico
Difesa
capacità di modulare le attività delle cellule nervose
regolazione della concentrazione degli ioni
smaltimento dei mediatori chimici
Le cellule della nevroglia si diversificano per morfologia, funzione, origine e sono:
derivate dal neuroectoderma (spongioblasti):
astrociti
oligodendrociti
cellule ependimali
derivate dai monociti del sangue:
cellule della microglia
derivate dalle creste gangliari:
cellule di Schwann
le cellule satelliti
Astrociti
sono le cellule più numerose nell’ambito della nevroglia e contengono abbondanti strutture filamentose dette gliofibrille, scomponibili in gliofilamenti di 100-130 A° di diametro appartenenti alla categoria dei filamenti intermedi e costituiti da una proteina acida detta proteina acida gliofibrillare (GFAP).
Sono distinguibili 2 tipi morfologici:
astrociti fibrosi
sono più abbondanti nella sostanza bianca
presentano dei prolungamenti più sottili e più lunghi che a volte assumono l’aspetto di lamine che si insinuano negli spazi esistenti tra le altre cellule
contengono abbondanti gliofibrille
astrociti protoplasmatici
sono più abbondanti nella sostanza grigia
hanno prolungamenti più corti e grossolani (tozzi) dei fibrosi
sono meno ricchi di gliofilamenti
Possiedonoi molti organuli simili ai lisosomi, cioè vescicole rotondeggianti molto elettrondense (gliosomi)
funzioni degli astrociti:
a)sostegno meccanico e rivestimento
gli astrociti di entrambi i tipi si mettono in rapporto gli uni con gli altri con i loro prolungamenti (anche mediante nexus) formando una rete che accoglie i corpi cellulari, i dendriti e le prime porzioni degli assoni delle cellule nervose.
alcuni astrociti, più vicini alla superficie del sistema nervoso centrale e in particolare dell’encefalo, mandano dei prolungamenti in direzione della superficie esterna del sistema nervoso centrale dove si slargano e diventano simili ad un epitelio appiattito, saldano i propri margini e formano una specie di rivestimento nei confronti del tessuto sottostante (membrana limitante gliale)
b) sostegno trofico e barriera emato-encefalica
gli astrociti più vicini ai vasi sanguigni inviano alcuni loro prolungamenti ai vasi stessi avvolgendoli:le sostanze nutritizie, per raggiungere le cellule nervose, devono attraversare la parete dei vasi e le strutture di rivestimento formate dagli astrociti (barriera ematoencefalica)
l’esistenza di questa barriera è dimostrata dal fatto che molti farmaci, anche se iniettati in circolo (endovena) non possono raggiungere le cellule nervose.
il ruolo preponderante nella costituzione della barriera è però svolto dalle cellule endoteliali: infatti le cellule endoteliali dei capillari delle cellule nervose presentano i loro margini strettamente legati da specializzazioni del tipo della zonula occludens mentre nella maggior parte dell’organismo le cellule endoteliali lasciano fra i loro margini delle fenestrature
Gli astrociti contribuiscono alla barriera endoteliale producendo fattori della matrice extracellulare (glicoproteine: aggrina ecc) che inducono le caratteristiche tipiche di queste cellule endoteliali
c) cicatrici gliali
gli astrociti entrano in gioco quando sopravvengono dei fenomeni che portano a distruzione il tessuto nervoso (processi infiammatori, emorragie, ischemie o neoplasie cerebrali)
quando il tessuto nervoso si è distrutto le cellule nervose non possono rigenerarsi (?) e lo spazio vuoto viene occupato da proliferazione degli astrociti: si costituisce una cicatrice gliale
d) Ruolo nella trasmissione sinaptica
Recentemente si è visto che le cellule astrocitarie possono regolare la concentrazione degli ioni K+ negli spazi sinaptici e possono riciclare mediatori chimici
Oligodendrociti
Sono le cellule già studiate nella costituzione della fibra mielinica della sostanza bianca.
Cellule ependimali
epitelio cubico/cilindrico le cui cellule possono essere più o meno alte (il loro aspetto ricorda allora un epitelio cilindrico pseudostratificato; tappezza la cavità interna del sistema nervoso centrale
nel versante libero le cellule ependimali presentano dei microvilli piuttosto irregolari e in certi casi una o più ciglia vibratile.
la parte prossimale (che è quella verso il tessuto nervoso circostante) presenta (specialmente nel periodo embrionale) un lungo prolungamento detto fibra ependimale.
questo lungo prolungamento può attraversare lo strato del tessuto nervoso e raggiungerne la superficie esterna, a questo livello si comporta come i prolungamenti degli astrociti, costituendo con essi la membrana limitante gliale.
ruolo delle cellule ependimali nella produzione del liquor
al livello dei ventricoli cerebrali, le cellule ependimali, oltre che formare il rivestimento più interno del sistema nervoso, entrano nella costituzione dei cosiddetti plessi corioidei.
infatti in prossimità della volta dei ventricoli accade che, durante lo sviluppo embrionale, non si verifica proliferazione di cellule nervose
in conseguenza di questo fatto la meninge più interna (Pia Madre ricca in vasi sanguigni) si viene a trovare in stretta vicinanza con le cellule ependimali
i capillari derivati da questi vasi , in queste sedi, diventano più abbondanti e formano dei ciuffi di anse capillari che si spingono all’interno delle cavità ventricolari rimanendo rivestiti da uno strato di cellule ependimali
queste strutture formate da capillari sanguigni rivestiti dall’ ependima sono i plessi corioidei e nel loro insieme costituiscono la tela corioidea
il plasma sanguigno, passando attraverso la parete del capillare e le cellule ependimali, modifica la concentrazione in proteine, sali e diventa liquido cefalo-rachidiano o liquor
quindi i plessi corioidei sono la sede di produzione del liquido cefalorachidiano e le cellule ependimali con molta probabilità partecipano alla produzione del liquor stesso
questo è dimostrato dal fatto che le cellule ependimali dei plessi corioidei sono ricche di mitocondri e di strutture microvillari
Il liquor prodotto a livello dei ventricoli cerebrali circola nelle cavità interne del sistema nervoso e a livello del quarto ventricolo, mediante i forami di comunicazione, può portarsi dalla cavità interna allo spazio subaracnoideo, compreso cioè tra pia madre e aracnoide
all’interno dello spazio subaracnoideo Il liquor costituisce una specie di cuscinetto idraulico che serve a proteggere il tessuto nervoso.
Cellule satelliti
derivano dalle creste gangliari e si dispongono attorno ai corpi cellulari dei neuroni gangliari
sono molto sottili e non sempre evidenti nelle sezioni dove lasciano semmai intravedere il loro nucleo
Cellule della microglia
sono cellule con origine completamente diversa rispetto alle altre perché derivano dai monociti del sangue e raggiungono il sistema nervoso proprio quando si verifica l’inizio della vascolarizzazione
come altre cellule derivate dai monociti svolgono funzioni fagocitarie.
si tratta di cellule dotate di un piccolo corpo cellulare contenente il nucleo e di alcuni prolungamenti che si ramificano notevolmente fra gli spazi compresi fra le altre cellule
per la loro attività fagocitaria intervengono ogni volta che nel sistema venoso si verificano dei processi distruttivi, infiammatori, necrotici, emorragici (spazzini del sistema nervoso)
come i macrofagi del connettivo, hanno capacità di presentare l’antigene e quindi sono responsabili di processi immunologici
fonte: http://www.med.unipi.it/morfologia/istologia/istolembmed/SMOTlez/08%20Tessuto%20nervoso.doc
Autore del testo: non indicato nel documento di origine
Tessuto Nervoso
Il sistema nervoso è organizzato anatomicamente in:
- sistema nervoso centrale (SNC) comprende il cervello e il midollo spinale
- sistema nervoso periferico (SNP) comprende i nervi cranici che derivano dal cervello e i nervi spinali emergenti dal midollo spinale con i gangli.
Dal punto di vista funzionale si divide in:
- componente sensitiva (afferente) che riceve e trasmette impulsi al SNC
- componente motoria (efferente) che si origina dal SNC che trasmette impulsi in tutto il corpo. Questa è divisa in:
- sistema somatico - gli impulsi che si originano dal SNC vengono direttamente trasmessi per mezzo di un singolo neurone, alla muscolatura scheletrica.
- sistema autonomo – gli impulsi vengono trasmessi ad un ganglio per mezzo di un neurone e successivamente da un altro neurone del ganglio si origina un impulso che raggiunge la muscolatura liscia, miocardica e le ghiandole.
Oltre ai neuroni nel tessuto nervoso troviamo anche altre cellule che fanno parte delle cellule di nevroglia, che non trasmettono impulsi ma hanno una funzione di supporto.
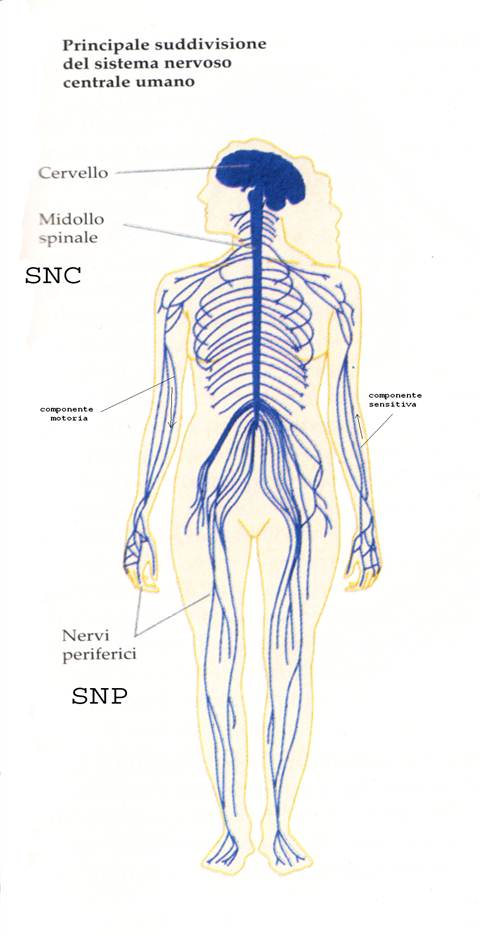
Gangli
si definisce ganglio nervoso un insieme di corpi cellulari situato esternamente al SNC. Si tratta di piccole strutture ovoidali incapsulate da tessuto connettivo denso. In base alla morfologia e alla funzione si possono distinguere due tipi di gangli nervosi:
gangli cerebrospinali (sensitivi)
gangli autonomi (associati ai nervi del sistema autonomo)
Sistema nervoso centrale
Il SNC è composto da sostanza grigia da sostanza bianca.
La sostanza bianca è costituita da fibre mieliniche, oligodendrociti, astrociti fibrosi e cellule di microglia. Il colore bianco è dato dalla mielina.
La sostanza grigia contiene il soma (corpo cellulare), fibre amieliniche e mieliniche, astrociti protoplasmatici, oligodendrociti e cellule di microglia.
Nelle sezioni trasverse di midollo spinale la sostanza bianca è localizzata all’esterno e la sostanza grigia all’interno, ove assume una forma ad H. Nel tratto centrale dell’H si trova una cavità, il canale centrale, residuo del tubo neurale tappezzato di cellule ependimali. La sostanza grigia forma le corna anteriori dell’H contenente neuroni motori dai quali si originano le radici ventrali dei nervi spinali. Anche le corna dorsali dell’H sono di sostanza grigia che ricevono fibre sensitive dai neuroni dei gangli spinali.
Il SNC è protetto dal cranio e dalla colonna vertebrale e inoltre da membrane di tessuto connettivo dette meningi. Dalla più esterna le meningi sono:
- Dura madre
- Aracnoide
- Pia madre
La pia madre e l’aracnoide sono connesse e si considera come un'unica membrana detta pia aracnoide.
Sistema nervoso della vita di relazione comprende centri e fibre sensitive (ganglisensitivi) e centri e fibre motrici (motilità volontaria)
Sistema nervoso autonomo comprende centri e fibre sensitive e centri e fibre motrici (simpatico e parasimpatico)
I neuroni
Sono le cellule responsabili della ricezione e della trasmissione degli impulsi nervosi da e verso il SNC.
I neuroni possono essere divisi in tre zone:
1.un corpo cellulare
2.dei prolungamenti detti dendriti
3.un unico prolungamento detto neurite o assone
corpo cellulare
è detto anche pirenoforo o soma o anche perikarion. Esso rappresenta la parte centrale della cellula dove è presente il nucleo ed il citoplasma. Il citoplasma è ricco di reticolo endoplasmatico rugoso ricco di cisterne e poliribosomi. Colorati opportunamente (coloranti basici) appaiono al microscopio come corpi di Nissl. I mitocondri hanno creste longitudinali invece che trasversali. L’apparato di Golgi, vicino al nucleo è ben sviluppato in relazione alla produzione di neurotrasmettitori.
I neuroni del SNC si distinguono da quelli del SNP per la forma:
Neuroni del SNC: sono poligonali con numerosi processi citoplasmatici
Neuroni del SNP: sono rotondeggianti con un unico prolungamento citoplasmatico.
Dendriti
Dal pirenoforo si dipartono i dendriti che svolgono la funzione di ricezione di stimoli sensoriali provenienti da cellule, assoni e neuroni. La modalità di arborizzazione dei dendriti è particolare per ogni tipo di assone. La porzione terminale delle ramificazioni dei dendriti permette la formazione di sinapsi che consentono la ricezione di molti impulsi e la trasmissione di segnali ad altri dendriti.
Neurite o assone
Dal lato opposto dei dendriti si origina il neurite o assone, prolungamento citoplasmatico privo di reticolo endoplasmatico rugoso e ricco di mitocondri, che può raggiungere il metro di lunghezza. Esso conduce l’impulso allontanandolo dal pirenoforo verso altri neuroni, cellule muscolari e ghiandole. La parte terminale si arborizza secondo terminazioni dette bulbi terminali o bottoni terminali, quali si appoggiano ad altre cellule formando le sinapsi, regioni ove si trasmette l’impulso nervoso alle cellule. Il diametro dell’assone è proporzionale alla velocità di trasmissione del segnale. Si possono distinguere alcune parti dell’assone:
- segmento iniziale: porzione compresa fra il suo punto di partenza e l’inizio della guaina mielinica che lo avvolge.
- Guaina mielinica: gli assoni possono avere una guaina mielinica e sono detti assoni mielinizzati, in essi l’impulso viaggia più velocemente. La guaina mielinica può essere assente attorno all’assone formando gli assoni amielinici. La guaina mielinica deriva dalle cellule della nevroglia che depositano la guaina attorno all’assone. L’assone mielinizzato ha un aspetto biancastro. Questa caratteristica permette di distinguere nel SNC la sostanza bianca da quella grigia.
Classificazione dei neuroni
I neuroni sono classificati in quattro tipi sulla base della loro forma:
-
neuroni unipolari (possiedono un unico prolungamento e sono molto rari nei vertebrati)
-
neuroni bipolari (presentano un singolo assone e un singolo dendrite. Si trovano nell’epitelio olfattivo della mucosa nasale )
-
neuroni pseudounipolari (presentano un unico prolungamento che parte dal soma, dopo un breve tratto si biforca in due rami disposti a T uno che entra nel SNC e l’altro che raggiunge la periferia.
-
neuroni multipolari (dotati di più prolungamenti uno dei quali è l’assone e gli altri i dendriti).
Possono essere classificati anche sulla base della loro funzione:
-
neuroni sensitivi (afferenti) – sono specializzati nella ricezione di impulsi sensoriali sulla loro terminazione dendritica e a trasmetterli al SNC per la elaborazione
-
neuroni motori o motoneuroni (efferenti) si originano dal SNC e portano gli impulsi ai vari organi e cellule, muscolari, ghiandolari e altre cellule nervose.
-
Interneuroni – si trovano nel SNC e hanno la funzione di collegare e di integrare le cellule nervose sensitive e motorie per formare una rete di circuiti nervosi. Il loro numero è stato elevato dall’evoluzione del sistema nervoso.
Cellule di nevroglia o gliali
La funzione metabolica e di supporto dei neuroni è svolta dalle cellule di nevroglia anche dette cellule gliali. Sono in grado di recuperare gli ioni e i prodotti del metabolismo dei neuroni, come il potassio il glutammato e altro che si accumula attorno ai neuroni. Partecipano al metabolismo energetico dei neuroni liberando glucosio dai loro depositi di glicogeno. Gli astrociti delle zone periferiche del SNC formano uno strato cellulare continuo attorno ai vasi sanguigni costituendo probabilmente la barriera emato-encefalica.(si ipotizza che questa in effetti derivi dai complessi di giunzione presenti negli endoteli cerebrali).
Le fibre nervose consistono di assoni neuronali avvolti da particolari guaine di origine ectodermica. Gruppi di fibre nervose costituiscono i fasci dell’encefalo e dl midollo spinale e i nervi periferici. Si incontrano differenze nelle guaine che avvolgono gli assoni a seconda che le fibre facciano parte del SNC o del SNP.
Nel tessuto nervoso adulto la maggior parte degli assoni è avvolta da pieghe singole o multiple di una cellula di rivestimento inguainante, rappresentata dalla cellula di Schwann nelle fibre del SNP e dall’oligodendrocito nelle fibre dl SNC, in ogni caso da cellule gliali o della nevroglia.
Il numero delle cellule di nevroglia è 10 volte più alto rispetto a quello dei neuroni. –Conservano la capacità di dividersi per tutta la vita. Non sono coinvolte nella conduzione nervosa. Si dividono in cellule localizzate nel SNC e in quelle localizzate nel SNP:
SNC – astrociti, oligodendrociti (che formano la macroglia), la microglia e le cellule ependimali.
SNP - cellule di Schwann
Astrociti (SNC)
Si conoscono due tipi di astrociti:
astrociti protoplasmatici – presenti nella sostanza grigia del SNC
astrociti fibrosi – presenti nella sostanza bianca del SNC
I due tipi cellulari potrebbero rappresentare una risposta funzionale di un'unica cellula sottoposta a condizioni ambientali diverse. Nel loro interno sono presenti fasci di filamenti intermedi costituiti da una proteina specifica per queste cellule, detta proteina acida fibrillare della glia.
astrociti protoplasmatici – sono cellule a forma stellata con molto citoplasma nucleo abbondante e diversi prolungamenti citoplasmatici ramificati. L’estremità di alcuni prolungamenti termina con un pedicello o piede vascolare che forma dei contatti con i vasi sanguigni. Altri nelle parti superficiali del cervello o del midollo spinale presentano dei prolungamenti a forma di pedicello che prendono contatto con la pia madre, costituendo la membrana pia-gliale.
Oligodendrociti (SNC)
Sono simili ai dendrociti, ma più piccoli e con meno prolungamenti. Sono presenti sia nella sostanza grigia che in quella bianca.
Si distinguono due tipi:
oligodendrociti interfascicolari- presenti fra i fasci di assoni, responsabili della formazione e del mantenimento della guaina mielinica attorno agli assoni. Sono simili alle cellule di Schwann, ma mentre quest’ultime sono in grado di avvolgere un singolo assone, gli oligodendriciti avvolgono più assoni contemporaneamente.
Oligodendrociti satelliti – sono strettamente adese al corpo cellulare dell’assone. La loro funzione non è nota.
Cellule ependimali (SNC)
Derivano dal rivestimento interno del tubo neurale e formano un epitelio cubico o cilindrico ciliato alle volte, con la funzione di muovere il liquido cerebrospinale. Rivestono la cavità dei ventricoli cerebrali ed il canale del midollo spinale. Alcune di loro si modificano nei ventricoli partecipando alla formazione dei plessi corioidei, responsabili della formazione del liquido cerebrospinale.
La microglia (SNC)
Il corpo cellulare è piccolo, di forma ellittica, il nucleo ha forma allungata con l’asse maggiore parallelo a quello del corpo cellulare. Si riconoscono poiché le altre cellule hanno nuclei tondi. Possiedono prolungamenti brevi ramificati. Alcune di loro hanno capacità fagocitaria e costituiscono il sistema fagocitario del tessuto nervoso.
Cellule di Schwann (SNP)
Si avvolgono attorno agli assoni nel SNP, formando il rivestimento mielinico. Sono appiattite con nucleo piatto, pochi mitocondri e un piccolo apparato di Golgi. La mielina è costituita dal plasmalemma della cellula che si avvolge più volte attorno all’assone. Ad intervalli regolari la guaina si interrompe e queste regioni amieliniche si indicano come nodi di Ranvier.
Il segmento di fibra compreso fra due nodi di Ranvier successivi si dice internodo o segmento internodale, esso è occupato da una sola cellula di Schwann.
Nella mielina è possibile osservare al M.E. delle fessure a forma di imbuto dette incisure di Schmidt-Lantermann, tali fessure contengono citoplasma della cellula di Schwann.
La sinapsi e la conduzione dll’impulso nervoso
Le sinapsi sono siti dove gli impulsi nervosi passano da una cellula presinaptica (neurone) ad un’altra cellula postsinaptica (un neurone, una cellula muscolare o ghiandolare).
Le sinapsi quindi permettono la comunicazione fra neuroni e fra questi e le cellule effettrici.
La trasmissione dell’impulso nervoso può avvenire o elettricamente o chimicamente
Riconosciamo quindi due tipi di sinapsi:
Sinapsi elettriche
Sinapsi chimiche
Sinapsi elettriche
Sono poco frequenti nei mammiferi, si incontrano nella retina e nella corteccia celebrale. Sono realizzate tramite giunzioni comunicanti o nexus, che permettono libero flusso di ioni da una cellula all’altra. Quando si realizza fra neuroni si genera flusso dicorrente. La trasmissione dell’impulso è più veloce nelle sinapsi elettriche.
Sinapsi chimiche
Rappresentano il modo più frequente di comunicazione fra due cellule nervose. La membrana presinaptica libera uno o più neurotrasmettitori nelle fessure intersinaptiche , spazi fra la membrana presinaptica della prima cellula e la membrana postsinaptica della seconda cellula.
Il neurotrasmettitore diffonde attraverso lo spazio intersinaptico e si lega ai recettori della membrana postsinaptica.
Il legame sui recettori scatena l’apertura dei canali ionici che consentono il passaggio di ioni che modificano la permeabilità della membrana postsinaptica ed invertono il potenziale di membrana.
Quando lo stimolo sulla sinapsi porta la depolarizzazione della membrana postsinaptica ad un livello tale da provocare un potenziale d’azione, si parla di potenziale postsinaptico eccitatorio.
Quando al contrario uno stimolo della sinapsi porta ad un aumento della polarizzazione si crea un potenziale postsinaptico inibitorio.
Le sinapsi chimiche possono essere divise in:
- sinapsi assodendritiche (fra un assone e un dendrite)
- sinapsi assomatiche (fra un assone e un soma)
- sinapsi assoassoniche (fra due assoni)
- sinapsi dendrodendritiche (fra due dendriti)
colonna vertebrale
Fonte: http://www.uniroma2.it/didattica/BIOLAPP/deposito/Tessuto_Nervoso.doc
sito web: http://www.uniroma2.it/didattica/
Autore del testo: non indicato nel documento di origine
Tessuto nervoso riassunto
- il tessuto nervoso
INTRODUZIONE
Il tessuto nervoso è formato da una rete di cellule comunicanti tra loro che sono le unità strutturali e funzionali del sistema nervoso. Questo sistema è diffuso in tutto l’organismo.
Le principali funzioni del tessuto nervoso sono:
- la comunicazione
- la secrezione endocrina.
Per quanto riguarda a comunicazione:
- il tessuto nervoso mette in comunicazione tutte le parti dell’organismo e l’ambiente esterno
- riceve stimoli dall’interno e dall’esterno ed è in grado di rielaborarli e stimolare risposte appropriate e coordinate tra i vari organi
La funzione endocrina è attuata da cellule nervose che in partocolari zone producono e rilasciano ormoni.
Il sistema nervoso è la sede dei fenomeni psichici.
CLASSIFICAZIONE DEL SISTEMA NERVOSO.
Negli umani il sistema nervoso viene distinto in due componenti con funzione differente:
- sistema nervoso cerebro-spinale o somatico, detto anche della vita di relazione
- sistema nervoso autonomo o simpatico, detto anche della vita vegetativa.
Il sistema nervoso cerebro-spinale, controlla la muscolatura volontaria ed è formato da due tipi di sistemi tra loro collegati:
- sistema nervoso centrale (SNC): costituito dall’encefalo (cervello+cervelletto) e da parte del midollo spinale.
- Sistema nervoso periferico (SNP): costituito dai nervi (cranici, spinali, ramificazioni e gangli)
Le cellule nervose si situano:
- nell’SNC
- nei gangli
I gangli sono complessi formati da neuroni con altre cellule satelliti, che fungono da supporto.
I nervi sono i prolungamenti delle cellule nervose sottoforma di fibre, che hanno conduzione centrifuga o centripeta.
Il sistema nervoso autonomo o simpatico controlla le contrazioni involontarie della muscolatura liscia ed è formato da:
- sistema nervoso simpatico: situato in posizione tronco-lombare, che causa reazioni di eccitazione
- sistema nervoso ortoimpatico: è situato nella zona cranio-sacrale, ha funzioni maggiormente inibitorie.
Gli organi nervosi sono formati da differenti tessuti:
- tessuto nervoso
- tessuto della nevroglia
- tessuto vascolare.
Le cellule della glia e il microcircuito sono di fondamentale appoggio ai neuroni, cosicché gli organi nervosi sono dei veri e propri complessi pluritissutali.
IL NEURONE
Il sistema nervoso è formato da due tipi di cellule:
- i neuroni,
- le cellule della glia, che hanno funzioni di sostegno.
Il neurone è l’unità fondamentale e funzionale per l’espletamento della mansione deputata al sistema nervoso:
- la ricezione, la conduzione e la trasmissione dell’impulso.
Le principali caratteristiche attribuite ai neuroni sono:
- irritabilità: capacità di reagire agli stimoli e di trasformarli in impulsi nervosi
- conducibilità: capacità di trasmettere gli stimoli
- polarizzazione funzionale: l’impulso si propaga secondo direzioni preferenziali
L’impulso nervoso è permesso dal passaggio di ioni Na+ e K+ nell’ambiente intra o extracellulare in modo da creare una differenza di potenziale:
- la differenza di potenziale che si trasmette causa il rilascio di neurotrasmettitori a livello delle sinapsi, giunzioni particolari tipiche del tessuto nervoso.
- Vengono trasmessi alle cellule effettrici o ad altri neuroni.
Le componenti morfologiche principali di u neurone sono:
- il pirenoforo: detto anche corpo o soma, rappresenta il corpo cellulare del neurone
- i dendriti: sono prolungamenti citoplasmatici solitamente ramificati e numerosi.
- Il neurite o assone: è un unico lungo prolungamento citoplasmatico
- La fibra nervosa: è un prolungamento degli assoni rivestita da involucri. Può avere una distanza notevole, fino a superare il metro di lunghezza.
All’inizio del Novecento, Camillo Golgi scoprì il metodo di indagine dell’impregnazione argentica, che consentiva di osservare un numero limitato di neuroni:
- Cajal, riuscì a scoprire la dottrina del neurone, in cui affermava che questi formano reti complesse tra loro rimanendo unità indipendenti e giungendosi attraverso particolari giunzioni, chiamate sinapsi.
- Con lo sviluppo delle tecniche, si riuscì a seguire le molecole che fluivano negli assoni e a determinare i circuiti che vi sono in essi.
- Con l’impiego di tecniche immunocitochimiche ed immunoistochimiche si è riuscito a comprendere le modalità con cui il neurone svolge la propria funzione.
Infine si è potuto dimostrare che il neurone non è solamente una pura unità morfologica:
- esso è un’unità secernente, capace di elaborare secreti specifici, attivi, di molteplice natura e funzione
- si è così dimostrata l’esistenza di un sistema endoneuronale secernente.
In definitiva, si è storicamente ampliato il concetto di neurosecrezione, che ora intende una modalità operativa con cui i neuroni comunicano tra loro e con le cellule effettrici, secernendo:
- messaggeri chimici specifici con meccanismo neurocrino: i neurotrasmettitori e le proteine di adesione.
- Ormoni, con vere e proprie funzionalità endocrine, detti neurormoni,
- proteine sinaptiche, con funzionalità autocrine.
NUMERO E FORMA DEI NEURONI
Secondo la concezione di Bizozzero i neuroni sono elementi perenni:
- si sviluppano con fenomeni mitotici molto presto durante il differenziamento embrionale
- giungono nell’adulto ad un numero approssimativo di 1011,
Nonostante si fosse creduto che i neuroni fossero elementi sostanzialmente immobili, oggi si deve prendere un concetto come vero, la neuroplasticità:
- i neuroni possono modificare alcune caratteristiche morfofunzionali (forma, dimensioni, struttura citoscheletrici e giunzioni sinaptiche) in relazione a sollecitazioni di varia natura
- anche in fase di riparazione si può osservare la neuroplasticità.
Le forme sono molto eterogenee, ma la più classica è quella di un pirenoforo triangolare a cui si legano i vari prolungamenti dendritici.
LA MEMBRANA DEL NEURONE
La membrana del neurone, costituisce lo strato limite periferico che circonda il neurone e i suoi prolungamenti dendritici e assonico.
In corrispondenza delle sinapsi, la membrana assume strutture particolari.
La membrana è la responsabile della trasmissione dell’impulso intraneuronale.
Nella membrana sono presenti tre tipi di proteine intrinseche degne di nota:
- proteine canali
- proteine pompa
- proteine recettori
Talvolta le funzioni di un complesso proteico possono anche essere unificate: ad esempio un recettore può essere legato ad una proteina canale.
Le proteine recettoriali, sono siti scaglionati nella membrana pre e postsinaptca:
- ancorano neurotrasmettitori e ormoni.
Le proteine canale attraversano tutta la lunghezza degli assoni e le membrane pre e postsinaptiche:
- consentono il passaggio di ioni Na+, K+, Ca2+ e Cl-
- sono qualificate come canali ionici e attivate da stimoli di natura differente.
Le proteine canale ioniche sono costituite da:
- più subunità
- ogni subunità è formata da 6 catene ad alfa-elica
- ogni elica possiede delle regioni ad ansa che sporgono verso l’esterno e verso l’interno della membrana con funzione di riconoscimento e trasduzione
Secondo la natura degli stimoli che possono attivare le proteine canale si possono distinguere:
- canali ionici voltaggio-dipendenti: l’apertura e la chiusura dei canali è regolata da differenze di potenziale che si instaurano lungo la membrana dell’assone
- canali ionici attivati da ligando chimici: possiedono un sito recettoriale che aggancia dei neurotrasmettitori o degli ormoni
- canali ionici attivati da pressione o stiramento: indotto da filamenti citoscheletrici che ancorano la proteina canale in corrispondenza del poro sul versante endocellulare.
La membrana assonico è polarizzata:
- vi è una differenza di potenziale di -70 mV tra l’interno dell’assone e l’esterno
- La concentrazione di ioni K+ è dieci volte maggiore all’interno rispetto all’esterno
- La concentrazione di ioni Na+ è circa 10 volte maggiore all’esterno.
- La differenza di potenziale è mantenuta da una proteina ATPasica detta pompa sodio/potassio, che pompa il sodio all’esterno e il potassio all’interno dell’assone.
La trasmissione dell’impulso lungo l’assone è data da un’oda di depolarizzazione che si innesta a livello del pirenoforo:
- a livello del pirenoforo si abbassa la differenza di potenziale e si ha l’apertura dei canali del sodio
- il sodio fluisce all’interno abbassando il potenziale. Si ha una doppia reazione, ovvero quella di aprire i canali del sodio successivi e di aprire quelli del potassio in loco, facendo ristabilire localmente la ddp con la fuoriuscita di potassio
- alle spalle del fronte dell’onda di depolarizzazione le pompe sodio/potassio ristabiliscono la situazione originaria e impediscono la propagazione retrograda dell’impulso.
Nelle fibre amieliniche il processo di depolarizzazione è piuttosto lento, poiché l’attivazione consequenziale delle pompe e l’espulsione degli ioni comporta spreco di tempo e di energia:
- le velocità di propagazione è di circa 1-2 m/s.
Nelle fibre mieliniche le proteine canale sono disposte solamente a livello dei nodi di Ranvier:
- si ha una propagazione dell’impulso con conduzione saltatoria, poiché le aperture dei canali avvengono discontinuamente solo a livello dei nodi di Ranvier
- questo permette anche risparmio energetico, poiché vi è meno sodio da riportare all’esterno e meno potassio da reintrnalizzare da parte delle pompe
- la velocità di conduzione dell’impulso è molto più alta, fino a 120 m/s, e varia in funzione alla larghezza della guaina mielinica.
In sintesi, le proteine canali voltaggio-dipendenti e le proteine pompa sono le responsabili della conduzione dell’impulso nervoso intracellulare.
IL PIRENOFORO.
Il pirenoforo è il centro trofico de neurone, ed è costituito da un citoplasma, detto pericarion che circonda il nucleo, in cui sono immersi i vari organuli:
- RER
- Mitocondri
- Complesso di Golgi
- Lisosomi
All’interno del pirenoforo, si osservano due componenti che sono caratteristiche dei neuroni:
- sostanza tigroide
- neurofibrille
Il nucleo ha notevoli dimensioni e forma sferica:
- presenta cromatina finemente dispersa (eucromatina)
- è poco colorabile
- è solitamente unico e disposto al centro (ad eccezione delle cellule di Purkinje che sono tetraploidi e le gangliari simpatiche che possono essere binucleate)
- il nucleolo è ben evidente e colorabile al centro del nucleo
La sostanza tigroide è una formazione morfologica visibile in seguito a colorazione con coloranti basici:
- è molto presente nei grandi neuroni,
- poco presente nei piccoli neuroni sensitivi
- è costituita da cisterne di RER unite a numerosi poliribosomi liberi
Il RER, oltre ad essere presente nel pericarion sotto forma di sostanza cromofila (tigroide) è anche presente nei dendriti di grandi dimensioni, ma è assente negli assoni.
Il REL è abbondante:
- si estende anche nei dendriti e nell’assone
- appare sottoforma di tubuli e vescicole di profilo irregolare.
Il complesso di Golgi è presente in tutte le cellule nervose:
- molto sviluppato
- costituito da pile di cisterne appiattite attorno al nucleo
I mitocondri sono molto frequenti e di piccole dimensioni:
- hanno forma a bastoncello
- si interpongono tra le varie cisterne
- sono molto presenti lungo i dendriti e gli assoni disponendosi in maniera ordinata.
- Contengono enzimi legati ai processi di ossidoriduzione e altri processi indispensabili per la vita del neurone
I lisosomi sono ben presenti in tutti i tipi di neuroni:
- svolgono le funzioni correlate al ricambio degli organuli citoplasmatici
- contengono sostanze litiche per degradare materiale di rifiuto
In tutto il pirenoforo si possono osservare delle reti di filamenti sottili e numerosi, dette neurofibrille:
- sono aggregati di neurofilamenti, filamenti intermedi di diametro di circa 10 nm costituiti da differenti tipi di proteine
- sono strutture che dal pirenoforo decorrono anche lungo gli assoni e i dendriti
- terminano a livello delle sinapsi
- sono molto abbondanti negli assoni di grosso calibro.
Oltre alle neurofibrille formate da neurofilamenti, sono presenti anche dei microtubuli (neurotubuli) che si estendono con regolarità all’interno del neurone:
- sono osservati in quantità maggiore negli assoni di piccolo calibro
- sono frequenti nei dendriti.
I neurotubuli e i neurofilamenti sono presenti in quantità differenti a seconda del tipo di neurone, tuttavia, sono maggiori i neurofilamenti negli assoni e maggiori i neurotubuli nei dendriti.
I microfilamenti, sono associati a numerose proteine e svolgono le seguenti funzioni:
- stabilizzano le proteine transmembrana fornendogli un sito di attacco
- regolano il posizionamento delle vescicole presinaptiche
- sono responsabili della mobilità dei coni di crescita e del posizionamento assonico.
- Intervengono nella rigenerazione delle fibre nervose.
DENDRITI
I dendriti sono prolungamenti citoplasmatici che si staccano, solitamente in un numero elevato, da pericarion e sono rivestiti da membrana plasmatica.
Sono prolungamenti di calibro ampio e possono presentare ramificazioni con un angolo inferiore a 90°.
I dendriti sono prolungamenti relativamente corti, la cui lunghezza non supera mai i 700 µm, si assottigliano man mano che si allontanano dal corpo cellulare:
- non posseggono rivestimenti particolari (al contrario degli assoni, es. la guaina mielinica)
- la struttura e le ramificazioni dei dendriti caratterizzano i vari tipi di neuroni.
La struttura citoplasmatica dei dendriti varia in funzione del loro calibro e della distanza dal pirenoforo:
- la composizione è essenzialmente uguale a quella del citoplasma
- compaiono vescicole di REL
- scompare quasi totalmente il RER con l’aumentare della distanza dal pericarion
- i mitocondri sono orientati e presentano una periodicità nella lunghezza del dendrite.
- Sono ricchi di microtubuli che si orientano paralleli all’asse dendritico.
La superficie dei dendriti appare irregolare ed è ricoperta da molte estroflessioni ovoidali, dette spine:
- sono di dimensioni e altezza variabile da 1 a 5 µm
- sono costituite da un collo e da un bulbo allargato con forma ovoidale
- sono zone di contatto sinaptico con gli assoni di altre cellule nervose.
- Non sono presenti strutture citoplasmatiche all’interno, ma si ha solamente l’apparato della spina, costituito da tre o quattro cisterne allargate e con disposizione parallela.
La superficie dei dendriti rappresenta, con le sue numerose giunzioni sinaptiche con altri neuroni (sinapsi asso-dendritiche) una struttura fondamentale per un neurone nell’interazione con informazioni provenienti da più fonti.
In determinate condizioni, i dendriti sono anche in grado di trasmettere degli impulsi oltre che riceverli:
- si stabiliscono delle sinapsi dendro-dendritiche che possono influenzare neuroni adiacenti
Il numero dei dendriti varia con l’età, ovvero diminuisce con l’invecchiamento:
- si pensa che possano essere la struttura per cui si abbia l’apprendimento e la memoria
- è stato dimostrato che la fosfatidilserina riduce la perdita delle spine dendritiche indotta dall’invecchiamento cerebrale.
ASSONE O NEURITE
L’assone o neurite è il prolungamento del neurone che conduce gli impulsi dal pirenoforo a distanze anche considerevoli:
- ha un notevole sviluppo in lunghezza
- la larghezza varia da 1 µm a 20 µm e influisce sulla velocità di trasmissione dell’impulso.
- Terminano con formazioni specializzate dette bottoni terminali sinaptici.
Gli assoni possono presentare un rivestimento quale:
- la guaina mielinica
- il neurilemma.
Ogni cellula ha un unico neurone:
- generalmente diparte dal pirenoforo
- può accadere che inizi da un dendrite
L’assone ha un contorno liscio e può, ad una certa distanza dal nucleo, emettere delle diramazioni che si assottigliano:
- le diramazioni permettono ad un neurone di influire su più cellule
Gli impulsi nervosi percorrono l’assone in direzione centrifuga. L’impulso può essere condotto tramite:
- sinapsi: giunge e si collega ad un altro neurone
- giunzioni citoneurali: si può connettere con ghiandole, cellule muscolari, fibre muscolari striate, fibrocellule lisce, ecc…
La membrana dell’assone è denominata assolemma.
Il citoplasma dell’assone è denominato assoplasma e include:
- mitocondri
- neurofilamenti
- microtubuli
- microfilamenti di actina
- vescicole delimitate da membrane lisce.
I neurofilamenti sono la componente principale del citoscheletro dell’assoplasma, ma in prossimità delle ramificazioni i microtubuli diventano la componente principale.
Non sono presenti anche numerosi filamenti di actina.
Il REL è scarso e disorganizzato, ma sono presenti vecicole ricoperte da membrane lisce che si situano tra i microtubuli e i microfilamenti.
Il RER è del tutto assente.
I mitocondri appaiono sottili e allineati orizzontalmente, in maniera parallela all’asse del neurite:
- sono più frequenti in zone presinaptiche.
Il punto in cui l’assone emerge dal pirenoforo è detto cono di emergenza:
- presenta microtubuli con un aspetto fascicolato
- è sprovvisto di sostanza tigroide.
Il primo tratto dell’assone, in cui non è ancora ben presente la guaina mielinica, prende il nome di segmento iniziale:
- ha una soglia di eccitabilità inferiore al pirenoforo
- presenta uno strato di materiale elettrodenso che è riscontrabile anche in prossimità dei nodi di Ranvier.
FLUSSO ASSONICO.
Al’interno degli assoni, il citoscheletro ha differenti elementi con specifiche funzioni:
- neurofilamenti: funzione meccanica di sostegno
- microtubuli: hanno la funzione di trasporto
Nell’assone, lungo i microtubuli vi è un flusso bidirezionale di vescicole:
- le sostanze che l’assone utilizza alla sua terminazione gli devono giungere da pirenoforo
- il materiale endocitato e le membrane da riciclare ritornano tramite un flusso anterogrado al pirenoforo.
Si è potuto dimostrare che esistono due livelli di velocità a seconda del materiale da trasportare. Si individuano quindi:
- flusso assonico lento
- flusso assonico veloce.
Il flusso assonico rapido è in grado di trasportare vescicole con una velocità costante di 50-400 mm al giorno.
Questo flusso rapido coinvolge il trasporto di:
- vescicole contenenti neurotrasmettitori chimici
- sostanze implicate nella funzione neurosecretoria
- costituenti di ricambio della membrana plasmatica.
Il flusso rapido è bidirezionali e comprende:
- movimento anterogrado: avviene dal pirenoforo alla terminazione dell’assone. Vede il trasporto di vescicole con neurotrasmettitori e organuli
- movimento retrogrado: avviene nel senso opposto. Riporta al pirenoforo le membrane da riciclare e organuli destinati ad essere degradati dai lisosomi.
Il flusso assonico lento si attua in due differenti velocità:
- 5 mm al giorno: riguarda componenti citosolici come la clatrina, l’actina, la calmodulina, ecc..
- 1 mm al giorno: è il trasporto che comprende il riciclaggio dei materiali del citoscheletro.
Le modalità con cui avviene il trasporto assonico rapido sono legate all’azione di specifiche proteine motrici con attività ATPasica:
- il trasporto anterogrado avviene grazie alla chinesina, che possiede due catene avvolte con due teste in cui si lega ATP e si legano i microtubuli
- il trasporto retrogrado avviene grazie ad una classe di dineine che, sempre con attività e teste ATPasiche, permettono il movimento in senso inverso sui microtubuli.
Per quanto riguarda le modalità del trasporto assonico lento non si hanno dei dati sperimentali precisi.
CLASSIFICAZIONE DEI NEURONI.
Le principali tipizzazioni dei neuroni sono fatte:
- in base al comportamento dell’assone
- in base alle modalità delle ramificazioni dei prolungamenti e alla direzione dell’impulso.
In base al comportamento dell’assone si possono distinguere:
- neuroni del 1° tipo di Golgi: l’assone è relativamente lungo e mantiene la sua unicità a notevole distanza dal pirenoforo.
- Neuroni del 2° tipo di Golgi: con un neurite corto, che a breve distanza si ramifica in maniera complessa, perdendo molto presto la propria individualità.
In base alle modalità di ramificazione dei prolungamenti, al loro numero, alla direzione degli impulsi, con una classificazione più recente, si posson distinguere:
- neuroni unipolari: sono i neuroni embrionali e i neuroni sensitivi. Presentano un solo prolungamento che trasmette l’impulso in maniera centrifuga.
- Neuroni bipolari: sono forniti di entrambi i tipi di prolungamenti, un assone e un solo dendrite che si generano ai poli opposti del pirenoforo
- Neuroni pseudounipolari: sono neuroni che sembrano forniti da un solo tipo di prolungamento, ma in realtà i due prolungamenti sono confluiti in un solo ramo che fuoriesce dal pirenoforo e si apre a T per dividersi in due prolungamenti. Sono i neuroni dei gangli cerebrospinali.
- Neuroni multipolari: hanno un assone e molti dendriti. Sono i neuroni più frequenti.
FIBRE NERVOSE
Molto spesso gli assoni sono rivestiti da guaine che li avvolgono per quasi tutta la lunghezza:
- la fibra nervosa è l’insieme morfofunzionale dell’assone e del suo rivestimento
- collegano le varie parti del sistema nervoso e conducono gli stimoli
- sono strutture filamentose che possono raggiungere anche notevoli lunghezze.
CLASSIFICAZIONE DELLE FIBRE NERVOSE
L’assone, in base al suo tipo di rivestimento forma differenti tipologie di fibre nervose:
- fibre mieliniche: il mesassone delle cellule gliali si avvolge ripetutamente sul neurite
- fibre amieliniche: non si ha un rivestimento costante da parte delle cellule della glia
- assone nudo: non presenta alcun tipo di rivestimento.
FIBRE NERVOSE MIELINICHE.
Le fibre nervose mieliniche si trovano sia nel sistema nervoso periferico che in quello centrale.
Le fibre nervose mieliniche del SNP presentano una guaina mielinica fornita dalle cellule di Schwann e formano i nervi di:
- nervi spinali
- nervi encefalici
Le fibre nervose mieliniche del SNC hanno una guaina costituita dagli oligodendrociti, cellule che, a differenza delle cellule di Schwann avvolgono più di un assone. Queste fibre nervose costituiscono:
- sostanza banca
- radici dei nervi periferici.
Le cellule gliali che avvolgono gli assoni formando fibre nervose presentano un citoplasma scarso, che si riscontra nelle porzioni esterne e in prossimità dei nodi di Ranvier.
A differenza delle fibre nervose del SNP, quelle del sistema nervoso centrale non sono circondate da tessuto connettivo, ma sono in rapporto con altre cellule della neuroglia.
FIBRE NERVOSE AMIELINICHE
Le fibre nervose amieliniche sono fibre nervose prove di guaina mielinica.
Nel sistema nervoso periferico sono circondate soltanto dal citoplasma della cellula di Schwann:
- sono assenti i nodi di Ranvier
- le cellule di Schwann formano un rivestimento continuo lungo l’assone
- appaiono pallide o grige poiché non hanno il classico colore della mielina
Nel sistema nervoso centrale si trovano numerose fibre amieliniche, che però mancano della guaina di Schwann (o neurilemma):
- sono assoni privi di avvolgimenti specifici
- sono in rapporto con le cellule gliali circostanti formando un fitto intreccio.
L’assone si presenta nudo all’origine dell’assone e a livello delle terminazioni.
NEURILEMMA
Nelle fibre del sistema nervoso periferico le cellule di Schwann forniscono il rivestimento dell’assone e insieme alla loro lamina basale costituiscono la guaina di Schwann o neurilemma:
- nelle fibre mieliniche il neurilemma è posto al di sopra della guaina mielinica
- nelle fibre amieliniche la guaina di Schwann riveste direttamente l’assone.
Le cellule di Schwannsono le principali cellule non eccitabili del tessuto nervoso periferico:
- avvolgono in cilindri l’assone,
- si dispongono a file allineate con regolarità
- ogni segmento di fibra in cui vi è una cellula di Schwann si chiama segmento internodale
- il tratto di fibra tra un internodo e l’altro si chiama nodo di Ranvier.
GUAINA MIELINICA
Le fibre mieliniche appaiono al microscopio ottico come tubuli omogenei, traslucidi e rifrangenti:
- la guaina mielinica conferisce colore e lucentezza caratteristici alla sostanza bianca dell’encefalo e nel midollo spinale
- la guaina si colora in maniera differente a seconda delle colorazioni che si attuano.
Una guaina mielinica riveste la maggior parte delle fibre del SNC e del SNP.
La mielina rappresenta un ottimo isolante elettrico:
- l velocità di trasmissione degli impulsi nervosi aumenta con l’aumentare del diametro della fibra
La guaina mielinica riveste gli assoni in maniera discontinua, formata da una successione di segmenti internodali:
- tra due segmenti internodali sono presenti delle strozzature che formano i nodi di Ranvier.
- A livello dei nodi, l’assone è rivestito solamente dalle cellule di Schwann e dalla loro membrana basale.
Nelle fibre del SNP la guaina mielinica presenta incisore a forma di imbuto tra le lamelle disposte a distanze regolari, chiamate incisure di Schmidt-Lantermann.
La guaina mielinica è costituita da strati concentrici di lamelle lipoproteiche che presentano una alternanza di linee scure e chiare secondo un modello periodico.
FORMAZIONE DELLA GUAINA MIELINICA
La mielina è costituita dalla membrana plasmatica della cellula di Schwann o degli oligodendrociti.
Il processo di avvolgimento della membrana plasmatica segue le seguenti modalità:
- inizialmente l’assone è circondato dalla cellula di Schwann,
- le estremità della cellula di Schwann si incontrano e formano il mesassone.
- Il mesassone si allunga e si avvolge a spirale sull’assone
- Man mano si ha l’avvolgimento il citoplasma viene espulso
- L’assone si trova formato da due anelli addossati di membrana plasmatica
- L’avvolgimento continua fino a numerosi giri, in cui la membrana plasmatica si giustappone continuamente.
Una cellula di Schwann, avvolgendosi forma un segmento internodale o internodo di mielina:
- il nucleo rimane situato, insieme ad una porzione di citoplasma, all’esterno della fibra
- il nucleo si pone circa a metà dell’internodo
La regione in cui le estremità dei processi delle cellule di Schwann si unisce ad una porzione della membrana della cellula stessa è denominata mesassone.
Le fibre mieliniche sono quindi formate da un avvolgimento di citoplasma delle cellule di Schwann, che può arrivare fino a 50 giri.
Le fibre amieliniche, invece, sono date dall’avvolgimento di più assoni da parte di una sola cellula di Schwann:
- in questo caso, il nucleo si pone pressoché centralmente e le porzioni citoplasmatiche formano solamente dei mesassoni con un'unica inclusione citoplasmatica.
- Anche nel caso delle fibre amieliniche, le cellule di Schwann si dispongono in serie.
NODI DI RANVIER
I nodi di Ranvier sono dei restringimenti periodici della guaina mielinica livello delle fibre nervose:
- l’avvolgimento elicoidale della mielina si interrompe
- si formano due cupole a livello degli avvolgimenti mielinici che formano delle sacche contenenti enzimi, vecsicole e microtubuli.
Nel nodo di Ranvier, la cellula di Schwann non si interrompe, ma va a ricoprire le parti in cui non v’è più mielina formando interdigitazioni con la cellula accanto:
- in quei bulbi che si vengono a creare, vi sono ampie porzioni di citoplasma delle cellule di Schwann, contenenti RER, ribosomi, glicogeno e microtubuli.
- Si forma un collare circolare, dato da una strozzatura della cellula di Schwann in prossimità dei nodi.
Sebbene la mielina, a livello dei nodi, si interrompe, l’assone non resta mai nudo:
- è rivestito da processi citoplasmatici delle cellule di Schwann che si interdigitano tra loro.
INCISURE DI SCHMIDT-LANTERMANN
Le incisure di Schmidt-Lantermann sono delle fessure imbutiformi che si presentano a livello internodale nella guaina mielinica:
- in queste tasche è presente un residuo di citoplasma della cellula di Schwann che ha formato la guaina mielinica
- questo residuo di citoplasma permette la comunicazione tra i vari strati della cellula
- le tasche che formano le incisure non contengono solitamente degli organuli
Al ME è visibile questa struttura in sezione longitudinale, ed appare come una incisura:
- in quella zona, la mielina resta lassa.
COMPOSIZIONE CHIMICA DELLA MIELINA
La mielina ha una composizione molto simile a quella delle altre membrane plasmatiche (lipidi e proteine), tuttavia presenta delle caratteristiche proprie.
Le percentuali in peso dei componenti sono:
- lipidi al 70 %
- proteine circa al 30 %
La componente lipidica presenta una particolarità data dalla presenza di sfingomieline e cerebrosidi.
Durante i tratti di bilayer fosfolipidico e sfingomielinico, non sono presenti proteine canale per ioni:
- la continuità dello strato lipidico permette un notevole isolamento elettrico
La componente proteica prevede:
- un gruppo di proteine basiche della mielina (MBP)
- una glicoproteina specifica associata alla mielina (MAG), con funzione di adesione e ancoraggio nel processo di mielinizzazione
Negli oligodendrociti, si può trovare prevalentemente una proteina caratteristica detta proteolipide (PLP).
Nelle fibre periferiche mature so trovano altre proteine di adesione che sono implicate nella mielinizzazione:
- P0 o MPZ
- PMP22, presente nella membrana delle cellule di Schwann.
Le alterazione dei geni che codificano per le proteine mieliniche portano a sindromi molto gravi, spesso con difficoltà deambulatorie.
NERVI PERIFERICI
Le fibre nervose, nel tratto che diparte dal corpo cellulare alle loro terminazioni in strutture periferiche, si raggruppano in fasci e formano i nervi periferici.
Le fibre connesse con il midollo spinale formano i nervi spinali, che fuoriescono dai dischi intervertebrali.
Le fibre nervose connesse con l’encefalo formano i nervi cranici, che escono dai fori situati alla base del cranio.
Nei nervi possono passare sia fibre efferenti, connesse a neuroni motori verso la periferia, sia fibre afferenti, che dai neuroni sensitivi si muovono verso i neuroni del sistema nervoso centrale.
ORGANIZZAZIONE DEI NERVI PERIFERICI.
In tutti i nervi, sia le fibre mieliniche che quelle amieliniche sono avvolte da una guaina esterna di tessuto connettivo denso detta epinevro.
Dall’epinevro dipartono setti connettivali che suddividono i fascetti di fibre nervose. Queste emanazioni di connettivo sono dette perinevro.
Le singole fibre, sono avvolte da connettivo lasso detto endonevro:
- forma una rete di fibre reticolari e materiale glicoproteico (proteoglicani e GAG) attorno alle cellule di Schwann, in continuità con la loro membrana basale
- nell’endonevro sono anche presenti numerosi capillari sanguiferi, che risultano ben isolati
Il liquido interstiziale dell’endonevro è isolato dal perinevro, che non ne permette la fuoriuscita.
Il perinevro forma attorno ai fascetti di fibre un manicotto di spessore variabile, formato da cellule appiattite unite da giunzioni strette (Tight junctions):
- all’esterno e all’interno del perinevro è presente una lamina basale che permette il passaggio di macromolecole.
GANGLI
I gangli sono raggruppamenti di cellule nervose disposte lungo il percorso dei nervi:
- i gangli sensitivi sono associati a nervi spinali e, più raramente, ai nervi cranici.
Ogni ganglio comprende:
- cellule nervose
- tessuto connettivo
- fibre nervose
- vasi sanguiferi
Il ganglio è delimitato da una capsula di connettivo denso che è in continuità con l’epinevro e il perinevro:
- trabecole di tessuto connettivo formano una rete in cui si inseriscono gli elementi cellulari, inframezzati dalle fibre nervose
- le cellule nervose sono ricoperte da cellule della glia dette cellule satelliti, che sono in stretta continuità con il tessuto connettivo
Le cellule nervose a livello dei gangli sono neuroni pseudounipolari:
- dal corpo cellulare diparte un solo prolungamento che si divide in due rami
- un ramo trasmette l’impulso dalla periferia al centro senza passare attraverso il corpo cellulare
- il corpo cellulare ha solamente funzione trofica.
SINAPSI
I neuroni, mantenendo la propria individualità morfologica, sono messi in contatto tra loro in modo da poter trasmettere e ricevere impulsi:
- ogni neurone ha la possibilità e la capacità di discriminare tra i vari impulsi e elaborare la risposta adeguata
Il contatto tra la terminazione assonica di un neurone e un altro neurone (a livello dei dendriti o del pirenofoto) prende il nome di sinapsi o giunzione sinaptica:
- la propagazione dell’impulso attraverso una sinapsi è unidirezionale
- un neurone può ricevere un numero variabile di terminazioni afferenti, a seconda della sua morfologia e localizzazione (si arriva a centinaia di migliaia nelle cellule di Purkinje)
Esistono due tipi di sinapsi lungo gli assoni:
- bottoni sinaptici terminali: è posto alla porzione terminale di un assone ed assume una forma grossolanamente tondeggiante.
- Boutons en passant: sono rigonfiamenti presenti lungo gli assoni. Normalmente sono presenti solo su fibre amieliniche, anche se è possibile riscontrarli a livello di alcuni nodi di Ranvier.
La fibra nervosa, a livello della giunzione sinaptica, perde il rivestimento mielinico e presenta una espansione che forma il bottone terminale.
La morfologia generale della sinapsi comprende tre zone:
- zona presinaptica: porzione terminale della cellula afferente, comprendente la membrana a contatto con l’elemento cellulare che riceve l’impulso e parte del citoplasma contenuto nel bottone
- zona postsinaptica: è la zona della cellula ricevente in cui si lega il bottone sinaptico dell’assone della cellula afferente.
- Fessura sinaptica: è lo spazio che intercorre tra le due membrane.
A livello dei bottoni, è possibile osservare numerose vescicole sinaptiche e un materiale elettrodenso adeso alle facce pre e postsinaptiche delle due membrane.
In base alle quantità del materiale elettrodenso si distinguono:
- sinapsi asimmetriche o di tipo I: il materiale adeso alla membrana postsinaptica è maggiore di quello della presinaptica. Si ritiene che siano sinapsi di tipo eccitatorio.
- Sinapsi simmetriche o di tipo II: la quantità di materiale elettrodenso è identica su entrambe le membrane. Si ritiene che siano di tipo inibitorio.
Le sinapsi sono di vario tipo anche a seconda degli elementi che si uniscono:
- asso-dendritiche: sono le sinapsi più comuni, possono essere asimmetriche o simmetriche
- asso-somatiche: si formano per la giunzione di assone e pirenoforo. Sono solitamente simmetriche
- sinapsi asso-spinose: si formano tra l’assone e le spine.
- Asso-assoniche
- Dendro-dendritiche: si ritiene possano essere reciproche, poiché si rinvengono polarità opposte alla stessa interfaccia giunzionale.
Altri tipi di giunzioni particolari esistono, ma sono poco frequenti.
ZONA PRESINAPTICA
La zona presinaptica è caratterizzata dalla presenza di:
- vescicole sinaptiche
- mitocondri
- corpi multivescicolari
- elementi di REL
Nelle vescicole, ricoperte da membrana unitaria, sono contenuti i mediatori chimici o neurotrasmettitori che vengono liberati all’arrivo dell’impulso nervoso:
- le dimensioni e il carattere elettrodenso della vescicola dipendono dal tipo di neurotrasmettitori che essa contiene
Nella maggior parte delle sinapsi del SNC e nelle giunzioni neuromuscolari, nelle vescicole sono contenuti:
- acetilcolina
- amminoacidi (glutammato, GABA, ecc…)
Altri bottoni terminali contengono una famiglia di neurotrasmettitori che comprendono:
- noradrenalina
- serotonina
- dopamina
La membrana presinaptica, presenta sul versante citoplasmatico una formazione geometrica regolare che viene indicata come griglia presinaptica:
- vi sono delle protuberanze che delimitano canali di circa 20 nm attraverso i quali le vescicole vengono direzionate nelle zone attive.
- Le vescicole riversano il materiale nella fessura sinaptica attraverso le zone attive per esocitosi
- La griglia presinaptica e le vescicole sono tutte in collegamento con una fitta rete di filamenti di actina.
FESSURA SINAPTICA
La fessura sinaptica è uno spazio di altezza 25-30 nm tra le membrane pre e post-sinaptiche:
- si nota la presenza di materiale filamentoso
- sono presenti molecole e matrice extracellulare coinvolte nella formazione e nella stabilizzazione della sinapsi.
ZONA POSTSINAPTICA
L’elemento strutturale che caratterizza funzionalmente la zona postsinaptica è la membrana postsinaptica:
- presenta numerose glicoproteine che fungono da recettori per i mediatori chimici che si riversano nella fessura sinaptica.
Al di sotto di tale membrana si trova un complesso elettrodenso e filamentoso che ha funzione strutturale detto apparato sottosinaptico.
VESCICOLE SINAPTICHE
Le vescicole sinaptiche sono il luogo di deposito temporaneo del neurotrasmettitori (mediatore chimico).
Le dimensioni e le caratteristiche di tali vescicole dipendono dal mediatore chimico in esse contenuto.
Si possono pertanto distinguere:
- piccole vescicole agranulari (SAV): contengono mediatori come l’acetilcolina
- piccole vescicole granulari (SGV): sono caratterizzate dalla presenza di un alone tra il core del granulo e la membrana limitante. Sono sede di accumulo di amine biogene, come ad esempio le catecolamine e la serotonina
- ampie vescicole granulari (LGV): hanno diametri compresi tra 100 e 200 nm. Non presentano alcun alone tra il core e la membrana limite. Contengono neurotrasmettitori purinici e peptidici.
MECCANISMO DI FUNZIONAMENTO DELLE SINAPSI.
Il segnale elettrico che raggiunge la terminazione presinaptica è convertito in segnale chimico:
- il neurotrasmettitori attraversa la fessura sinaptica e si lega al recettore sulla membrana postsinaptica
- a livello della membrana postsinaptica il segnale ricevuto dai recettori si riconverte in segnale elettrico
- si crea una onda di depolarizzazione a partire dalla membrana postsinaptica.
Quando in prossimità della membrana presinaptica arriva l’impulso elettrico, si aprono dei canali ionici che liberano ioni Ca2+, che sono i responsabili primari dell’ativazione del processo di esocitosi:
- la quantità di calcio liberata dipende dalla durata dell’impulso elettrico, che mantiene aperti i canali ionici
Le vescicole contengono una quantità costante di materiale da esocitare, cosicché, si riversa nella fessura sinaptica sempre una quantità discreta di neurotrasmettitori.
Le vescicole si dispongono in gruppi, i cui componenti vescicolari sono uniti da filamenti proteici.
La proteina che è implicata in queste connessioni è la sinapsina I:
- lega tra loro le vescicole
- lega i cluster vescicolari ad una proteina che emana dalla faccia citoplasmatica detta fondrina.
Prima del citoplasma è presente una rete di microfilamenti di actina che impediscono alle vescicole di fluire nelle zone attive.
Con l’arrivo dell’impulso nervoso la membrana presinaptica si depolarizza e si ha il rilascio massivo di ioni calcio:
- gli ioni Ca2+ si legano alla calmodulina, formando dei complessi che iniziano a fosforilare la sinapsina I.
- la sinapsina I perde affinità per le vescicole e per la fondrina e le rilascia
- le vescicole si portano verso le zone attive, ma sono trattenute dal reticolo di filamenti di actina
- la massiccia presenza di ioni calcio rende possibile l’attivazione delle actin-severing protein, che creano delle fessure nella rete di filamenti actinici.
- Le vescicole possono raggiungere la membrana ed essere esocitate.
La membrana della vescicola che in zona presinaptica si è fusa con la membrana dell’assone viene riassorbita per endocitosi e può essere riciclata, tramite il moto retrogrado.
A livello della membrana postsinaptica il neurotrasmettitori si lega a recettori che convertono il segnale chimico in segnale elettrico:
- i recettori sono canali ionici a controllo di ligando, che si attivano all’arrivo del neurotrasmettitori
- il rilascio di ioni permette di modificare il potenziale della membrana postsinaptica.
Le risposte di un neurone all’arrivo dell’impulso da parte di una sinapsi possono essere eccitatore o inibitorie, quindi si distingono:
- sinapsi eccitatore
- sinapsi inibitorie.
Nelle sinapsi eccitatorie il legame con il mediatore chimico determina l’apertura di canali con ioni positivi:
- si ha il rilascio di ioni positivi all’interno del citoplasma dendritico
- si crea un’onda di depolarizzazione nel neurone postsinaptico
- è il caso dell’acetilcolina per le giunzioni neuromuscolari.
Nelle sinapsi inibitorie invece si aprono canali di ioni negativi come quelli per Cl-:
- il potenziale di membrana viene mantenuto ad uno stato polarizzato o viene accentuata la polarizzazione opposta
- si impedisce l’insorgenza del potenziale d’azione
- si blocca l’impulso e il neurone rimane inibito.
È inoltre dimostrata l’esistenza a livello postsinaptico di secondi messaggeri di natura chimica, rilasciati con il legame di un neurotrasmettitore con il recettore:
- nucleotidi ciclici
- diacil-glicerolo
- fosfatidil-inositolo.
I mediatori chimici che si trovano all’interno della fessura sinaptica hanno vita breve, poiché una volta esaurita la loro funzione possono:
- essere degradati da enzimi litici
- essere riassorbiti dalla membrana presinaptica.
I mediatori chimici possono essere:
- acetil colina
- amine biogene (moradrenalina, istamina, dopamina, serotonina)
- amminoacidi (acido aspartico, acido glutammico, GABA)
- neuropeptidi
è ancora irrisolto il dubbio a proposito delle possibilità di conduzione di un solo tipo di neurotrasmettitore per ogni neurone:
- generalmente ogni neurone secerne solo un neurotrasmettitori
- è possibile che alcuni neuroni secernano anche un altro neuropeptide oltre al proprio mediatore specifico
Esistono anche delle sinapsi che non utilizzano mediatori chimici, ma conducono direttamente l’impulso elettrico. Sono membrane unite da giunzioni serrate per lo più sinapsi dendro-dendritiche:
- possono condurre l’impulso in due sensi
- la velocità di conduzione è maggiore
- sono scarsi nei mammiferi.
TERMINAZIONI NERVOSE SENSITIVE
Le terminazioni nervose sensitive possono:
- stabilire giunzioni sinaptiche con altri neuroni
- essere in rapporto con organi effettori
- ricevere stimoli che vengono condotti ai centri nervosi
Le vie afferenti del Sistema Nervoso sono connesse a un sistema di recettori specifici adibiti alla captazione di stimoli:
- esterocettori: captano gli stimoli dal mondo esterno
- enterocettori: captano gli stimoli all’interno dell’organismo
- propriocettori: ricevono gli stimoli dai muscoli, dai tendini e dalle articolazioni
La classificazione dei recettori in base al tipo si stimoli prevede:
- meccanorecettori
- chemiorecettori
- termorecettori
- fotorecettori.
Le modalità con cui le fibre terminano a livello dei recettori sono svariate e molto complesse.
Si possono distinguere le varie arborizzazioni dei terminali dei recettori in:
- terminazioni libere, le quali sono scoperte nei tessuti
- terminazioni sensitive capsulare, che hanno una capsula di speciale tessuto connettivo.
PLACCA MOTRICE
Le fibre muscolari scheletriche si contraggono in riposta a stimoli provenienti dal SNC, che giungono ai muscoli mediante l’assone di una fibra nervosa:
- una fibra nervosa serve numerose fibre muscolari
- l’insieme del neurone, dell’assone e delle fibre muscolari costituisce l’unità motoria.
Le ramificazioni della fibra nervosa terminano sulla fibra muscolare in punti specifici detti giunzioni neuromuscolari o placche motrici:
- l’assone perde la guaina mielinica
- la membrana basale dell’assone è in continuità con quella della fibra muscolare
- la regione priva di miofibrille in cui si innestano le ramificazioni è detta suola della placca motrice.
- Sono presenti numerosi nuclei, detti nuclei di arborizzazione che appartengono alle cellule di Schwann
- Il citoplasma della suola è ricco di mitocondri, RER e ribosomi liberi.
Nelle giunzioni neuromuscolari le fibre nervose terminano con una serie di espansioni bottoniformi, che si accolgono in invaginazioni del sarcolemma, dette docce sinaptiche:
- resta tra la membrana dell’assone e la fibra muscolare uno spazio di 25 nm
- in corrispondenza delle docce sinaptiche il sarcolemma si ripiega ulteriormente, formando delle fessure sinaptiche secondarie.
Le fessure secondarie si dispongono parallelamente tra loro, formando un sistema di pieghe:
- si forma un sistema di creste e gronde, con spessori della membrana differenti
- sono state individuate 5 proteine differenti associate alle creste e alle gronde
Associate alle creste si possono trovare recettori per l’acetilcolina e una actin-binding protein.
Associate alle gronde si riscontrano:
- anchirina
- proteina canale del sodio
- distrofia
In prossimità della membrana delle creste vi è uno spessore che, si suppone sia rappresentato da giunzioni aderenti con proteine come:
- vincolina
- talina
- alfa-actinina
- filamina
Nella terminazione presinaptica (dell’assone), si riscontrano:
- numerosi mitocondri
- vescicole contenenti acetilcolina.
La propagazione avviene nel modo seguente:
- il segnale elettrico proviene dal neurone e dalla doccia viene rilasciata acetilcolina
- questa si lega ai recettori specifici sul sarcolemma, determinando un aumento di permeabilità degli ioni
- nel sarcolemma si genera un’onda di depolarizzazione, che induce alla contrazione tutte le miofibrille.
L’azione della acetilcolina è brevissima:
- questa viene subito inattivata dall’enzima acetilcolinesterasi
- viene scissa la colina, che è poi recuperata dagli elementi presinaptica e riutilizzata per la sintesi di nuova acetilcolina.
TESSUTO GLIALE O NEVROGLIA.
INTRODUZIONE
La nevroglia o glia è un tessuto cellulare che rappresenta l’85% dei citotipi presenti nel tessuto nervoso:
- nonostante la maggioranza numerica, le cellule della glia sono considerate cellule di supporto ai neuroni
Queste cellule hanno, nel rendere il loro sostegno ai neuroni, funzioni molto importanti:
- funzione trofica
- funzione di riparazione
- protezione
- sostegno
La nevroglia è classificata in base al luogo in cu si situa a cui corrispondon citotipi differenti:
- del sistema nervoso centrale
- periferica
NEVROGLIA DEL SISTEMA NERVOSO CENTRALE.
All’interno del sistema nervoso centrale si presenta una glia composta da citotipi molto eterogenei, ognuno con una funzione differente.
Si riscontrano:
- ependimocito
- astrocito
- oligodendrociti
- microgliocito
Ciascuno di essi rappresenta un vero e proprio tessuto, tra cui si individuano:
- ependima
- macroglia
- oligodendroglia
- microglia
L’ependima costituisce il rivestimento delle cavità definitive scavate nello spessore del nevrasse:
- canale centrale del midollo spinale
- ventricoli cerebrali
L’ependima è costituito da un epitelio cubico-cilindrico monostratificato che riveste le cavità midollari e i ventricoli cerebrali:
- sono strettamente ravvicinati gli uni agli altri
- non presentano giunzioni occludenti, quindi è permesso il passaggio di macromolecole.
Un tipo di ependima modificato è quello delle cellule dei plessi coroidei, detti anche taniciti:
- sono complessi di cellule ripiegate su se stesse
- hanno funzione secernente (secernono il liquido cefalo-arachidonico)
- possiedono microvilli.
- Sono unite tra loro da giunzioni aderenti, quindi non lasciano passare i costituenti plasmatici attraverso l’endotelio fenestrato dei capillari coroidei
- I capillari coroidei fungono da membrana basale a queste cellule
La macroglia ha origine ectodermica ed è la forma di glia più distribuita nel nevrasse:
- le sue cellule sono gli astrociti, cellule dal corpo molto piccolo con molte espansioni citoplasmatiche ramificate che gli danno un aspetto stellato
- i bracci degli astrociti si collegano tra i pirenofori dei neuroni e i capillari dell’encefalo, poiché attuano scambi metabolici tra il flusso sanguigno e i neuroni
- sono le cellule gliali più numerose nella sostanza grigia (astrociti a brevi raggi).
L’oligodendroglia ha origine ectodermica e rappresenta il 40 % della glia del SNC:
- formata da oligodendrociti, che sono presenti sia nella sostanza grigia a contatto con i neuroni, sia in quella bianca a contatto con gli assoni
- gli oligodendrociti hanno funzione trofica nei confronti dei neuroni
- gli oligodendrociti sono i responsabili della mielinizzazione a livello del SNC. Una di queste cellule può rivestire più assoni
- presentano anche attività di sintesi di ormoni steroidei.
La microglia è formata da cellule che hanno origine mesenchimale e presentano esili espansioni citoplasmatiche che si connettono ai pirenofori a loro prossimi:
- la loro derivazione embrionale prevede la funzione di macrofagi, poiché giunti nel processo di vascolarizzazione del sistema nervoso
- la loro funzione è quella di ripulire e riparare le giunzioni assoniche o dendritiche lese.
GLIA DEL SISTEMA NERVOSO PERIFERICO
Nel sistema nervoso periferico la nevroglia è rappresentata da una notevole varietà di cellule:
- cellule capsulare o satelliti: avvolgono in modo più o meno completo i pirenofori dei neuroni pseudounipolari dei gangli sensitivi cerebrospinali e del sistema nervoso autonomo
- cellule di Schwann: mielinizzano assoni periferici
- cellule lemmali: formano il rivestimento capsulare dei recettori nervosi periferici
- cellule di teloglia: avviluppano le terminazioni nervose motrici
- cellule di sostegno: sono presenti insieme alle cellule sensitive secondarie nelle papille gustative.
BARRIERA EMATOENCEFALICA.
Il rapporto tra il tessuto nervoso e il sangue circolante è completamente differente da quello che si trova negli altri organi:
- vi è una barriera alla diffusione del sangue verso i neuroni
- la barriera è data dall’endotelio vasale e dalla sua membrana basale
I capillari del tessuto nervoso hanno un endotelio continuo, caratterizzato da:
- giunzioni strette: tight junctions formate da proteine di membrana quali l’occludina e la clausina.
- giunzioni aderenti: formate da caderine
E’ dimostrato che queste giunzioni giocano un ruolo importante nel realizzare una barriera funzionale che può ostacolare il passaggio di farmaci e batteri, come di altre macromolecole.
Si ritiene che gli astrociti, i quali espandono i loro terminali sui capillari, possano:
- indurre la formazione di giunzioni serrate.
- Interagire nella rimozione dell’eccesso di sodio nel microambiente perineuronale.
La barriera ematoencefalica è permeabile a:
- ossigeno
- anidride carbonic
- farmaci liposolubili
Tramite proteine recettoriali possono essere trasportati:
- glucosio
- nucleotidi
- vitamine
Tuttavia, la barriera ematoencefalica non è presente in tutte le zone cerebrali:
- è assente nelle zone circumventricolari (in prossimità delle ghiandole cerebrali come l’ipofisi) per permettere regolazioni endocrine.
Fonte: http://www.bluejayway.it/Enrico_Colombos_Page/Medicina_files/CORSO%20DI%20ISTOLOGIA.doc
Autore del testo: non indicato nel documento di origine
Tessuto nervoso riassunto
Visita la nostra pagina principale
Tessuto nervoso
Termini d' uso e privacy