Tessuto osseo
Tessuto osseo
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Le informazioni di medicina e salute contenute nel sito sono di natura generale ed a scopo puramente divulgativo e per questo motivo non possono sostituire in alcun caso il consiglio di un medico (ovvero un soggetto abilitato legalmente alla professione)
TESSUTO OSSEO
è un tipo particolare di tessuto connettivo in quanto è mineralizzato
questo gli conferisce particolari caratteristiche: è un tessuto resistente, ma presenta un certo grado di flessibilità e leggerezza
ha la funzione di:
sostegno: lo scheletro mantiene la posizione eretta
protezione: come altri tessuti connettivi svolgono una protezione che viene esplicata come rivestimento (ad esempio: scatola cranica, gabbia toracica)
movimento: nelle ossa si inseriscono i muscoli la cui contrazione è regolata dal sistema nervoso
metabolismo del Ca e P: il tessuto osseo ha grandi quantità di ioni (soprattutto calcio e fosforo):
non è un deposito stabile: a dispetto della sua apparente staticità, è un tessuto in continuo rimodellamento che produce movimento di ioni
emopoiesi: esiste il midollo osseo costituito da cellule emopoietiche parzialmente differenziate da cui prendono origine le cellule del sangue
composizione:
65% di cristalli di Ca sotto forma di cristalli di idrossiapatite
25% di proteine collageniche
2% di proteine non collageniche
10% di acqua
Il tessuto osseo può formare tipi diversi di ossa, cioè si organizza in modo da costituire:
osso compatto (ad occhio nudo non sono visibili delle cavità)
osso spugnoso (ad occhio nudo sono visibili delle cavità)
la struttura dell’osso è diversa, ma l’organizzazione microscopica del tessuto osseo è uguale in tutti e due i casi: si può parlare di lamelle ossee per entrambe le tipologie (organizzazione lamellare)
l’organizzazione non lamellare è presente nelle prime fasi di formazione dell’osso, nelle prime fasi di riparazione dell’osso, nei punti di inserzione dei tendini
ORGANIZZAZIONE GENERALE DELLE OSSA
classificazione delle ossa:
lunghe
corte
piatte
osso lungo: (esempio il femore) presenta un segmento centrale (diafisi) e due estremità (epifisi)
la diafisi è costituita da osso compatto che forma una parete che delimita la cavità midollare
le epifisi sono costituite da osso spugnoso rivestito da una lamina di osso compatto
l’osso spugnoso presenta delle cavità comunicanti tra loro e comunicanti anche con la cavità midollare della diafisi
l’osso compatto della diafisi si continua con l’osso compatto che riveste le epifisi
osso corto: (esempio il corpo vertebrale) ha struttura sovrapponibile a quella delle epifisi delle ossa lunghe
è costituito da osso spugnoso rivestito da una lamina di osso compatto
osso piatto: (esempio lo sterno) vi si distinguono due strati:
uno esterno e uno interno di osso compatto, che sono definiti tavolato esterno ed interno e che delimitano una parte centrale di osso spugnoso detta diploe
OSSO LAMELLARE
Le ossa sono per la massima parte rivestite sulla loro superficie esterna da periostio (strato connettivale denso all’esterno e connettivo reticolare con cellule osteogeniche a contatto con l’osso)
le cavità che si trovano all’interno delle ossa sono rivestite da endostio (costituito solo da strato di connettivo reticolare con cellule osteogeniche).
Nell’organismo adulto sia il tessuto osseo compatto che quello spugnoso sono costituiti da lamelle ossee, cioè da strati sovrapposti l’uno all’altro, ciascuno dei quali risulta costituito da cellule ossee disposte nella matrice ossea ricca di fibre collagene, sostanza amorfa (matrice organica) e sali minerali (matrice inorganica o mineralizzata)
ORGANIZZAZIONE DELLE LAMELLE OSSEE
Le lamelle ossee si possono disporre in varia maniera; ad esempio nelle ossa lunghe alcune lamelle (3-5) formano il sistema circonferenziale esterno che racchiude l’osso al di sotto del periostio.
A ridosso della cavità midollare si trovano altre 2-3 lamelle che formano il sistema circonferenziale interno;
tra questi due sistemi si trovano gli osteoni che somigliano a cilindretti solidi percorsi al loro interno in senso longitudinale da un lume cavo che contiene dei vasi. Le lamelle degli osteoni sono disposte concentricamente attorno al canale centrale. Gli osteoni si dicono anche sistemi di Havers e il canale centrale è denominato canale di Havers.
Si dicono invece canali di Volkman altri canali vascolari che decorrono perpendicolarmente rispetto all’asse maggiore degli osteoni.
Infine altre lamelle formano un sistema detto sistema interstiziale o breccia dell’osso che si trova tra gli osteoni; si tratta di osteoni in via di riassorbimento che assumono così una forma poliedrica per adattarsi alle superfici degli osteoni che stanno loro attorno.
COMPOSIZIONE DEL TESSUTO OSSEO
Le fibre collagene del tessuto osseo sono costituite da collagene di tipo I e sono molto numerose all’interno di ogni lamella.
La sostanza amorfa è poco rappresentata e costituisce l’1-2 % del peso secco dell’osso; contiene prevalentemente condroitinsolfati.
Sono presenti anche delle glicoproteine; oltre a quelle ricordate nel connettivo propriamente detto sono la osteonectina e la osteopontina.
La matrice inorganica o mineralizzata è costituita da cristalli aghiformi formati da sali di Ca (fosfati e carbonati) che nel complesso costituiscono la idrossiapatite.
Questi cristalli vengono posti in rapporto con le fibre collagene e si ritiene che la striatura tipica del collagene serva come stampo per la deposizione dei cristalli di idrossiapatite
CELLULE DEL TESSUTO OSSEO
Le cellule dell’osso sono:
le cellule osteoprgenitrici, gli osteoblasti, gli osteociti e gli osteoclasti
Cellule osteoprogenitrici:
si trovano nello strato interno del periostio e nell’endostio
somigliano alle cellule mesenchimali ed hanno capacità di differenziarsi in cellule osteoblasitche
possono anche dare origine a fibroblasti o a condrociti
Sono in genere di forma affusata e contengono uno scarso citoplasma debolmente basofilo
Osteoblasti:
sono le cellule responsabili della produzione dei costituenti dell’osso (fibre collagene e matrice organica) inoltre partecipano al meccanismo di mineralizzazione dell’osso.
Hanno forma rotondeggiante e posseggono brevi prolungamenti citoplasmatici; il loro citoplasma è intensamente basofilo e contiene anche dei corpi PAS positivi che sono stati messi in relazione con le sostanze enzimatiche implicate nella mineralizzazione della matrice.
Intervengono allorché si deve formare nuovo tessuto osseo: si riuniscono in piccoli gruppi attorno ai vasi e cominciano a produrre fibre collagene e sostanza amorfa (matrice osteoide).
A poco a poco il quantitativo della matrice osteoide aumenta e gli osteoblasti si dispongono in un’unico strato attorno ad essa.
Quando si vengono a creare le condizioni metaboliche ottimali per la mineralizzazione (precipitazione di sali di Ca) gli osteoblasti si dividono e formano uno strato di cellule più esterno alla prima fila di osteoblasti così continua la produzione di matrice osteoide, mentre lo strato di osteoblasti più vecchio (più interno) emette dei prolungamenti e si trasforma nelle cellule ossee mature cioè negli osteociti.
Questa divisione degli osteoblasti permette di continuare l’accrescimento dell’osso che altrimenti sarebbe interrotto quando le cellule vengono circondate dalla matrice mineralizzata.
Osteociti:
sono le cellule mature dell’osso;
sono contenuti in piccole escavazioni all’interno della matrice ossea dette lacune ossee;
il loro citoplasma è acidofilo.
Gli osteociti hanno forma molto irregolare essendo dotati di lunghi prolungamenti che sono circondati da matrice mineralizzata e sono contenuti nei cosiddetti canalicoli ossei.
Questi prolungamenti sono connessi alla loro terminazione con prolungamenti analoghi provenienti da cellule vicine.
La loro funzione è quella di connettere le cellule tra di loro e con i vasi che si trovano nel periostio, nell’endostio e nei canali vascolari (di Havers e di Volkmann).
Questa connessione rende possibile lo scambio di sostanze nutritizie e di segnali tra cellule incarcerate nella matrice mineralizzata.
Osteoclasti:
sono le cellule che provvedono al riassorbimento dell’osso che deve essere sostituito nel processo continuo di rimodellamento che ha luogo per tutto l’arco della vita.
Derivano dai monociti del sangue e sono in genere multinucleati tanto da somigliare a cellule giganti da corpo estraneo.
Hanno citoplasma acidofilo.
Sono accolti in lacune dette di Howship e con una loro superficie si mettono in rapporto con l’osso da riassorbire; a questo livello presentano un orletto striato.
Producono enzimi (fosfatasi acida e altri) e sostanze che permettono loro di distruggere la matrice che incontrano, di mobilizzare i sali minerali, riassorbire il calcio dalla matrice.
La loro funzione è controllata da ormoni (calcitonina e paratormone) e da vitamine (D)
Istogenesi dell’osso:
il tessuto osseo può costituirsi con due diverse modalità:
ossificazione membranosa: gli osteoblasti invadono il tessuto connettivo e iniziano la loro attività di produzione di matrice osteoide che successivamente va incontro a mineralizzazione. Si verifica ad esempio nelle ossa del cranio.
ossificazione condrale: consiste nella sostituzione di un modello di tessuto cartilagineo da parte di tessuto osseo neoformato. Si verifica ad esempio negli arti dove comunque il meccanismo condrale coopera anche con il meccanismo membranoso per formare le varie parti delle ossa lunghe (diafisi, epifisi, metafisi).
Fonte: http://www.med.unipi.it/morfologia/istologia/istolembmed/SMOTlez/06%20TESSUTO%20OSSEO.doc
Autore del testo: non indicato nel documento di origine
Tessuto osseo
Tessuto osseo e vibrazioni
In che modo stimoli meccanici sono in grado di modificare il livello di accrescimento dell’osso?
Il ciglio: un sensore per il mondo esterno
Quasi tutte le cellule di mammifero possiedono sulla loro superficie esterna un ciglio. Studi recenti hanno dimostrato che questo ciglio svolge una funzione di meccano sensore, di chemio sensore o di entrambe in base al tipo di cellula. Il cattivo funzionamento del ciglio comporta diverse patologie dell’uomo che hanno come sintomi disturbi alla visione, perdita di udito, polidattilia ecc... Le ultime ricerche hanno messo in luce il fatto che questi cigli rivestono un ruolo molto importante anche nello sviluppo e nell’omeostasi dello scheletro.
Il ciglio come meccano sensore nell’osso
Lo scheletro è la componente meccanica che ha la funzione di sostenere il corpo contro la forza gravitazionale e di aiutarlo nella locomozione. Lo scheletro si adatta agli stimoli meccanici: a stimoli meccanici maggiori corrisponde una sintesi maggiore di tessuto osseo; stimoli meccanici minori portano invece ad una minore sintesi e quindi ad una perdita di tessuto osseo.
L’osso è infatti un tessuto dinamico a lento turnover. Il processo di osteosintesi, sia nella fase di crescita sia nelle età successive, durante le quali si verifica un continuo rimaneggiamento dell’osso, ha come principale meccanismo di controllo lo stimolo meccanico costituto dalle tensioni e dalle deformazioni applicate all’osso sia dal carico sia dalla contrazione muscolare.
La capacità dell’osso di rispondere agli stimoli meccanici dipende delle cellule del tessuto osseo: osteoblasti, osteociti, osteoclasti. In particolare gli osteociti sono ritenuti i meccano sensori dell’osso a causa della loro posizione all’interno della matrice.
Essi infatti risiedono in delle cavità della matrice, dette lacune, da cui si dipartono esili canalicoli ramificati che contengono i prolungamenti citoplasmatici degli osteociti. Tramite il sistema di prolungamenti e dei canalicoli gli osteociti risultano essere in grado di effettuare scambi metabolici fra di loro e con il sistema circolatorio, scambi che sono favoriti dalla presenza di gap junctions tra le estremità dei prolungamenti degli osteociti. I primi a osservare la presenza del ciglio nell’osteocita furono Tonna e Lampen già nel lontano 1972 i quali ipotizzarono tra l’altro questo ciglio come il potenziale sensore per gli stimoli meccanici nell’osso. Il meccanismo attraverso il quale gli osteociti rispondono agli stimoli meccanici non è ancora del tutto chiaro, ma recenti esperienze hanno dimostrato che proprio questo ciglio è il sensore meccanico per il tessuto osseo. Questi cigli hanno infatti la capacità di captare gli stimoli meccanici provenienti dall’esterno, di discriminarli in base all’intensità, alla durata, all’orientamento e quindi di fare in modo che la cellula si comporti di conseguenza.
Esperienze e risultati
Recenti esperienze sono state fatte negli ultimi anni con lo scopo di capire che tipi di effetti hanno vibrazioni con caratteristiche definite (frequenza, intensità, durata...) sulle cellule dell’osso. In particolare nel 2006 un esperimento effettuato dal Dipartimento di Ingegneria Biomedica della State University di New York su un gruppo di femmine di topo BALB/cByJ di otto settimane ha dimostrato che l’applicazione di segnali meccanici di intensità estremamente bassa per un breve periodo di tempo al giorno (15 minuti al giorno) può attenuare già dopo tre settimane la perdita di tessuto osseo favorendone la formazione. Invece, segnali meccanici applicati per periodi di tempo più lunghi al giorno intervallati da brevi pause non hanno alcun risultato favorevole in merito. Successivamente studi preliminari su bambini disabili e donne nel periodo post-menopausale hanno dimostrato che questo tipo di segnali può essere molto efficace nella prevenzione dell’osso anche nella specie umana.
Due anni più tardi uno studio portato avanti sempre dal Dipartimento di Ingegneria Biomedica della State University di New York nel 2008 ha dimostrato che stimolazioni meccaniche di bassa intensità hanno effetti sulla proliferazione e sulla differenziazione delle cellule mesenchimale staminali promuovendo l’osteogenesi e prevenendo l’obesità. Le cellule staminali mesenchimali sono infatti caratterizzate dalla capacità di rinnovarsi e di differenziarsi in cellule che formano i tessuti come quello osseo e quello adiposo. Usando su maschi di topo C57BL/6J segnali meccanici di bassa intensità (0.2g, con una frequenza di 90Hz applicata per 15min al giorno, 5 giorni la settimana) la capacità di promuovere la produzione di tessuto osseo e di prevenire l’aumento del tessuto adiposo era collegato all’influenza degli stimoli meccanici sull’ambiente molecolare del midollo osseo. Sei mesi di queste vibrazioni hanno portato ad un aumento della produzione base di cellule staminali del 37% e delle cellule mesenchimali del 46%. Contemporaneamente all’aumento di cellule staminali è stato riscontrato un aumento della differenziazione osteoblastica contro quella adipocitica, aumento registrato dall’aumento della trascrizione del fattore Runx2 del 72% e dalla diminuzione del PPAR gamma del 27%. L’impatto fenotipico è risultato evidente dopo 14 settimane quando la formazione del tessuto adiposo è stata soppressa del 28%, mentre la frazione del volume dell’osso nella tibia era aumentato dell’11%.
Conclusioni
[L'osteoporosi è una patologia che attualmente colpisce dopo i 60 anni il 30 per cento delle donne e il 15 degli uomini. Durante questa patologia prevale la distruzione di sostanza ossea sulla neoproduzione di questa, con conseguente rarefazione progressiva del tessuto osseo. In genere si manifesta a livello di corpi vertebrali, ossa della mano e collo del femore.]
L’osso quindi risponde ad una grande varietà di segnali e la capacità dei segnali meccanici di influenzare la morfologia dell’osso dipende fortemente dalle sue caratteristiche e quindi dalla sua intensità, dalla sua frequenza e dalla sua durata..
Vibrazioni ad alta frequenza sono nello scheletro assile anabolici o anti-catabolici permettendo quindi una maggiore resistenza dell’osso. Lo sviluppo di concreti interventi biomedici nei campi dell’ortodonzia, della ricostruzione cranio facciale e per quanto riguarda l’osteoporosi richiederanno una precisa e dettagliata descrizione dei meccanismi di trasmissione di segnali meccanici nell’osso. Inoltre la modulazione meccanica della proliferazione e della differenziazione delle cellule staminali indica un importante programma terapeutico per promuovere la rigenerazione e la riparazione del tessuto osseo.
Autore: Lorenzo Cerrone
Fonte: http://www.salearning.org/focus/osseo.doc
Il tessuto Osseo
Estrarre pure da
http://www.atsaistudenti.it/articoli/il%20tessuto%20osseo.htm
Si parla di midollo che riempie le cavità dell’osso di cui quello giallo formato da grasso: è la parte che noi estraiamo con solvente per svuotare le cavità
Le cavità interne dell’osso sono occupate dal midollo osseo che può essere rosso, giallo oppure gelatinoso.
Il midollo rosso ha il compito di formare gli elementi figurati del sangue; il midollo giallo è prevalentemente formato da grasso e, in particolari circostanze, può trasformarsi in midollo rosso a funzione emopoietica;
Generalità
Il tessuto osseo e la cartilagine fanno parte dei tessuti connettivi specializzati per la funzione di sostegno della maggior parte degli organismi viventi. L'appartenenza del tessuto osseo ai tessuti connettivi è giustificata sia per la sua origine dal mesenchima, il tessuto embrionale che funge da matrice per tutti i tessuti connettivi, sia per la sua costituzione, essendo formato da cellule e da sostanza intercellulare composta da fibre collagene.
La peculiarità del tessuto osseo è quella di essere mineralizzato: infatti, la sostanza intercellulare è per la maggior parte impregnata di cristalli minerali, in prevalenza fosfato di calcio. La presenza di minerali, come pure l’abbondanza e la particolare distribuzione delle componenti organiche della sostanza intercellulare, conferiscono a questo tessuto proprietà meccaniche di durezza e di resistenza alla pressione, alla trazione e alla torsione. Oltre ad essere un tessuto ideale per il sostegno del corpo, rappresenta il principale deposito di ione calcio per le necessità metaboliche dell'intero organismo. La deposizione del calcio nell'osso e la sua mobilizzazione, controllate da meccanismi endocrini, contribuiscono in modo sostanziale alla regolazione dei livelli plasmatici di questo ione.
Da un punto di vista macroscopico, si distinguono due varietà di osso: l'osso spugnoso e l'osso compatto.
L'osso spugnoso si ritrova principalmente a livello delle ossa brevi, delle ossa piatte e delle epifisi delle ossa lunghe: ha questo nome in quanto appare conformato come una spugna, con travate ossee, dette trabecole, variamente orientate e intersecate tra loro e delimitanti cavità, dette cavità midollari, che in vivo sono ripiene di midollo osseo.
L'osso compatto forma la porzione più superficiale delle ossa brevi, delle ossa piatte e delle ossa lunghe, nonché a costituire la diafisi di queste ultime. Esso è privo di cavità macroscopicamente evidenti.
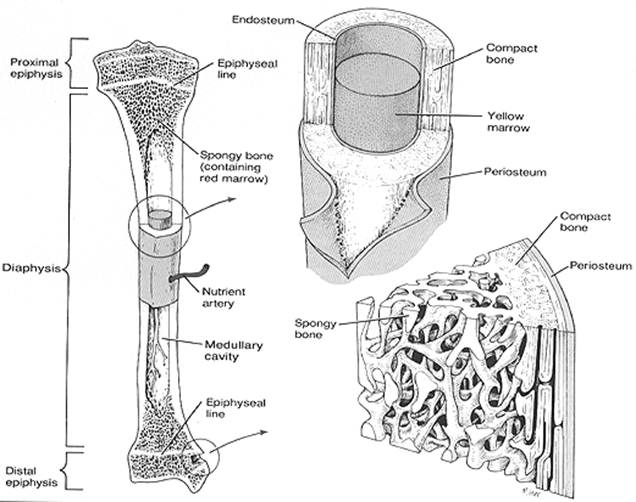
Questa immagine é stata scannerizzata dal libro di testo, “Basic Medical Anatomy”, di Alexander Spence (Benjamin/Cummings 1990).
Sostanza intercellulare del tessuto osseo
Facendo parte dei tessuti connettivi, il tessuto osseo contiene una quota rilevante di sostanza intercellulare organica, composta da fibre connettivali e da sostanza fondamentale anista, e di componenti minerali.
¨ Le fibre connettivali sono rappresentate per la quasi totalità da fibre collagene, composte da collagene di tipo I, che rappresenta il 90% del collagene totale ed entra a far parte dei tessuti connettivi, quali la pelle, i tendini, le ossa e la cornea.
Il collagene è composto da tre catene, avvolte insieme in una tripla elica stretta.
Questa struttura resistente è formata da una sequenza ripetuta di tre amminoacidi. Un amminoacido ogni tre è glicina, molecola di piccole dimensioni che si inserisce perfettamente nell'elica. Molte delle posizioni rimanenti nella catena sono occupate da altri due amminoacidi : prolina ed una sua versione modificata, l’idrossiprolina.
L'idrossiprolina, che è critica per la stabilità del collagene, viene sintetizzata modificando l'aminoacido prolina dopo che la catena di collagene è stata costruita. La reazione richiede la vitamina C per permettere l'addizione di ossigeno.
http://web.mit.edu/3.082/www/team1_f02/collagen.htm
Le fibre collagene sono molto resistenti, possono sopportare una trazione di 5 kg/cm2 e l'abbondante contenuto di collagene nell’osso è il principale responsabile della marcata acidofilia della sostanza intercellulare dell'osso.
¨ La sostanza fondamentale anista ha una composizione peculiare e in buona misura diversa da quella degli altri tessuti connettivi. Di essa fanno parte varie classi di macromolecole :
a) Il biglicano (PG-I) è costituito da due molecole di condroitinsolfato unite ad una estremità da un polipeptide ricco di leucina e lo si ritrova sia nella sostanza intercellulare mineralizzata che in quella non mineralizzata adiacente alle cellule ossee.
La decorina (PG-II) tende ad associarsi alle microfibrille collagene come a decorarle. E' formata da una parte proteica analoga a quella del PG-I unita ad una sola molecola di condroitinsolfato. La si ritrova nella sostanza intercellulare mineralizzata e si ipotizza che abbia un ruolo nell'orientare la deposizione dei cristalli minerali lungo le microfibrille collagene.
b) Le glicoproteine, di solito fosforilate o solfatate, includono molecole diverse alcune delle quali sono ritenute giocare un ruolo fondamentale nel controllo dei processi di mineralizzazione. Tra queste si annoverano:
- Osteonectina, la glicoproteina più abbondante, che ha un’alta affinità per il calcio, sia come ione libero che associato in complessi di tipo cristallino. Si ritiene che essa agisca come elemento di nucleazione dei cristalli minerali, in quanto ritenuta capace di concentrare il calcio nelle sue adiacenze creando così le condizioni per avviare la precipitazione del fosfato di calcio.
- Fosfatasi alcalina, unenzima capace di idrolizzare gruppi fosfato legati a substrati organici ed attivo in ambiente alcalino (pH 8-10). La sua funzione è oggetto di discussioni, in quanto potrebbe essere sia implicata nei processi di sintesi della matrice, sia come causa della formazione di cristalli.
Fibronectina, una molecola di adesione localizzata prevalentemente nella matrice pericellulare e caratterizzata da una porzione capace di legarsi al collagene. Si ritiene che la fibronectina sia coinvolta nei processi di migrazione, adesione alla matrice e organizzazione delle cellule dell'osso.
c) Le sialoproteine, o BSP (dall'acronimo inglese bone sialo-proteins), sono glicoproteine particolari contenenti residui glicidici di acido sialico. Queste proteine posseggono una sequenza aminoacidica particolare Arg-Gly-Asp (Arginina-Glicina-Acido aspartico) che in esperimenti in vitro è stata vista mediare l'adesione al substrato di svariati tipi cellulari, incluse le cellule dell'osso. Si ritiene pertanto che le sialoproteine ossee abbiano la funzione fisiologica di consentire l'adesione delle cellule alla matrice ossea.
d) Le proteine contenenti l'acido carbossiglutammico (GLA), che é un aminoacido particolare derivato dall'acido glutammico con un ulteriore gruppo carbossilico legato al carbonio. Il GLA incluso in una proteina possiede, nella porzione del residuo, due gruppi carbossilici liberi e ravvicinati che a pH fisiologico sono ionizzati e carichi negativamente, pertanto capaci di agire come una sorta di chelanti per i cationi bivalenti quali lo ione calcio. Le proteine dell'osso contenenti il GLA sono di due tipi.
Osteocalcina, o proteina GLA dell'osso,una piccola proteina contenente 3-5 residui di GLA. E' stato ipotizzato che essa possa giocare un ruolo di inibizione della mineralizzazione in quanto ritenuta capace di legarsi allo ione calcio e di renderlo indisponibile per la combinazione con lo ione fosfato, inibendo così l'accrescimento dimensionale dei cristalli minerali. Questa ipotesi è avvalorata dalla constatazione che l'osteocalcina abbonda nel tessuto osseo maturo ed è invece scarsa nel tessuto osseo in via di formazione, nonché dal reperto che questa proteina inibisce la crescita di cristalli di fosfato di calcio in vitro.
Proteina GLA della matrice, di peso molecolare maggiore della osteocalcina, è presente sia nell'osso maturo che in quello in via di formazione e anche nella cartilagine destinata a essere sostituita da tessuto osseo, come la cartilagine di accrescimento. Il suo ruolo biologico non è chiarito.
¨ La componente minerale è rappresentata da cristalli di sali di calcio, prevalentemente fosfato di calcio a cui si aggiungono quantità minori di carbonato di calcio e tracce di altri sali (fluoruro di calcio, fosfato di magnesio). Il fosfato di calcio è presente sotto forma di cristalli di apatite, la cui cella elementare ha la forma di un prisma esagonale appiattito e formula chimica Ca10(PO4)6++; le due cariche positive sono di norma neutralizzate dal legame con due ioni ossidrile (OH-), formando così la idrossiapatite, ma si possono ritrovare anche altri anioni (ione carbonato nella carbonatoapatite; ione fluoruro nella fluoroapatite). Il cristallo si origina dall'impilamento delle singole celle elementari ed ha la forma di un ago lungo e sottile, spesso circa 2 nm e lungo 20-40 nm.
I cristalli di apatite sono ben riconoscibili nei preparati di tessuto osseo allestiti per la microscopia elettronica in quanto fortemente elettrondensi; essi tendono a disporsi parallelamente tra sé e alle microfibrille collagene, di cui ricoprono la superficie e permeano le porosità. Osservazioni condotte durante il processo di mineralizzazione dell'osso hanno consentito di precisare che il fosfato di calcio precipita inizialmente sotto forma di minutissimi aggregati amorfi. Questi nuclei iniziali di concrezione minerale vengono rapidamente rimpiazzati da sottilissimi cristalli aghiformi disposti parallelamente a molecole filamentose della sostanza fondamentale detti filamenti assili (o crystal ghosts), verosimilmente costituiti da decorina, in rapporto col periodo delle microfibrille collagene. Tali cristalli crescono assumendo l'aspetto tipico dei cristalli di apatite, occupando progressivamente gran parte dello spazio interposto tra le microfibrille collagene e permeando le microfibrille stesse. Si ritiene verosimile che i precipitati di minerale fungano da centri di nucleazione dei cristalli di apatite, che una volta formatisi favoriscono la deposizione di nuovo minerale, che può avvenire sia per creazione di nuovi cristalli che per apposizione sui cristalli preesistenti. Tale fenomeno è finemente regolato dalle cellule ossee tramite la produzione di specifiche molecole della matrice ossea, come già accennato in precedenza. Secondo alcuni ricercatori la pirofosfatasi potrebbe attivarsi idrolizzando i pirofosfati organici presenti nell'osso. Tali molecole, quando sono integre, si combinano con le estremità dei cristalli di apatite e impediscono l'ulteriore concrezione di ioni calcio e fosfato, agendo quindi da inibitori della mineralizzazione. Quando vengono scissi dalla pirofosfatasi osteoblastica, i pirofosfati ossei perdono affinità per i cristalli di apatite consentendo così la crescita dei cristalli stessi.
Cellule del tessuto osseo
Le cellule proprie del tessuto osseo sono morfologicamente distinguibili in 4 varietà: le cellule osteoprogenitrici (dette anche preosteoblasti), gli osteoblasti, gli osteociti e gli osteoclasti.
¨ Le cellule osteoprogenitrici, o preosteoblasti, hanno forma fusata o ovalare, con citoplasma scarso. Esse si collocano sulle superfici libere delle ossa: le si riconoscono a livello dello strato più interno del periostio apposto all'osso, il cosiddetto strato osteogenico di Ollier, riccamente vascolarizzato. Tali cellule sono altresì localizzate a livello del tessuto connettivo lasso che riveste le cavita interne dell'osso, il cosiddetto endostio, in vicinanza dei capillari sanguigni. Le cellule osteoprogenitrici sono dotate di capacità proliferativa, che si manifesta in modo particolare durante l'accrescimento corporeo ma che può esplicarsi anche durante la vita adulta. Esse sono in grado di produrre e secernere le bone morphogenetic proteins (BMP), fattori di crescita e di differenziamento autocrini. Quando imboccano la via del differenziamento, le cellule osteoprogenitrici si trasformano in osteoblasti.
¨ Gli osteoblasti sono le cellule primariamente responsabili della sintesi della sostanza intercellulare dell'osso e della sua mineralizzazione. Esse hanno forma globosa o poliedrica e tendono a giustapporsi le une alle altre a formare delle lamine a ridosso delle superfici ossee in via di formazione. Gli osteoblasti hanno dimensioni rilevanti (circa 20 µm), un nucleo rotondeggiante ed un citoplasma abbondante. Istochimicamente, queste cellule si caratterizzano per la positività alla reazione per la fosfatasi alcalina. I mitocondri contengono numerosi granuli densi, interpretabili come accumuli di ioni calcio; sono visibili vescicole con vario aspetto contenenti molecole di procollagene; proteoglicani e altre molecole della sostanza fondamentale anista.. Gli osteoblasti sono uniti tra loro e con gli osteociti vicini tramite giunzioni serrate, attraverso le quali le cellule si scambiano molecole segnale per la coordinazione dell'attività metabolica e di deposizione della matrice ossea.
L'osteoblasto è la sede di sintesi delle molecole organiche della sostanza intercellulare dell'osso, le quali vengono successivamente esocitate ed assemblate all'esterno della cellula. L'osteoblasto presiede anche alla mineralizzazione della sostanza intercellulare, secondo modalità che non sono del tutto chiarite.
Gli osteoblasti sono coinvolti nei processi di rimaneggiamento dell'osso. Infatti, queste cellule sono in grado di innescare il riassorbimento della matrice ossea sia indirettamente, in quanto producono fattori solubili che attivano gli osteoclasti, le cellule preposte al riassorbimento osseo, sia direttamente, in quanto secernono enzimi capaci di scindere i componenti della matrice organica dell'osso.
¨ Gli osteociti sono le cellule tipiche dell'osso maturo, responsabili del suo mantenimento ed anche capaci di avviarne il rimaneggiamento. Sono cellule terminali, con una autonomia di vita finita, finemente regolata da meccanismi endocrini. Il corpo dell'osteocita rimane racchiuso in una nicchia scavata nella sostanza intercellulare ossea, detta lacuna ossea, la cui forma ricalca quella della cellula, mentre i prolungamenti sono accolti all'interno di sottili canali scavati nel tessuto osseo e definiti canalicoli ossei. Tra la membrana plasmatica del corpo cellulare e dei prolungamenti e la matrice mineralizzata rimane uno spazio occupato da tessuto osteoide che non mineralizza, attraverso il quale l'acqua e le sostanze disciolte (gas respiratori e metaboliti) riescono a raggiungere tutti gli osteociti, anche quelli più distanti dai vasi sanguigni. Metaboliti e molecole segnale disciolti nel citoplasma possono inoltre essere scambiate tra gli osteociti tramite delle giunzioni.
La mobilizzazione di ioni calcio dalla matrice ossea stimolata dal paratormone, un ormone che stimola il riassorbimento di calcio a livello renale, detto quindi ipercalcemizzate, è ritenuta dipendere principalmente dall'azione combinata di osteoblasti ed osteoclasti.
¨ Gli osteoclasti sono le cellule preposte al riassorbimento osseo. I precursori degli osteoclasti, detti preosteoclasti, nascono nel midollo osseo e sono apparentati con una categoria di globuli bianchi, i monociti. I preosteoclasti vengono trasportati dalla circolazione sanguigna fino alle sedi in cui devono avvenire processi di riassorbimento osseo; qui giunti, essi migrano nel tessuto osseo e si fondono insieme originando gli osteoclasti attivi, elementi capaci di dissolvere la componente minerale e di digerire enzimaticamente le componenti organiche del tessuto osseo.
Gli osteoclasti maturi sono cellule giganti (100-200 µm), plurinucleate in quanto originate dalla fusione dei singoli precursori mononucleati: in un singolo osteoclasto possono infatti essere presenti fino a 50 nuclei. L'osteoclasto attivato è aderente alla matrice mineralizzata in via di riassorbimento ed è solitamente accolto in una cavità, detta lacuna di Howship, che si forma a seguito dell'azione erosiva della cellula sull'osso. Una volta formata una prima lacuna, l'osteoclasto si distacca dalla matrice ossea, si muove su una porzione di osso adiacente a quella appena riassorbita, aderisce nuovamente e forma una nuova lacuna.
Organizzazione architetturale del tessuto osseo
In base alle dimensioni ed alla disposizione delle fibre collagene, si distinguono due varietà di tessuto ossseo, il fibroso e il lamellare.
¨ Il tessuto osseo fibroso è caratterizzato dalla presenza di fibre collagene di dimensioni rilevanti (5-10 µm di calibro). Il decorso di queste fibre non segue un orientamento definito, per cui esse appaiono intrecciarsi in tutte le direzioni dello spazio, così come avviene nei tessuti connettivi densi a fasci di fibre intrecciati. Il tessuto osseo fibroso è il primo ad essere deposto, sia durante lo sviluppo fisiologico che nella riparazione di fratture, dopo di ché esso viene rapidamente riassorbito e rimpiazzato con tessuto osseo di tipo lamellare. Ne rimane soltanto a livello delle inserzioni dei tendini e dei legamenti.
¨ Il tessuto osseo lamellare è la varietà più diffusa, costituisce la quasi totalità dell'osso compatto e buona parte dell'osso spugnoso. Esso è caratterizzato dalla ordinata disposizione delle fibre collagene e degli osteociti, che si dispongono in strati sovrapposti, detti lamelle ossee. A seconda della disposizione delle lamelle, si distinguono:
® Il tessuto osseo lamellare semplice, caratterizzato da un numero limitato di lamelle con andamento parallelo. Esso forma le trabecole e le lamine ossee più sottili, come le dialisi delle ossa lunghe.
® Il tessuto osseo lamellare osteonico, costituito da un numero variabile di lamelle (8-20) disposte concentricamente attorno ad un canale centrale che accoglie un vaso sanguigno, detto canale di Havers.
Nel tessuto osseo lamellare, le microfibrille collagene non tendono ad aggregarsi in fibrille né in fibre collagene. In ogni lamella le microfibrille collagene formano un feltro più o meno fitto intrecciandosi in ogni direzione dello spazio. A lamelle più sottili (circa 3 µm), ricche in microfibrille e relativamente povere in cristalli di apatite, si alternano lamelle più spesse (circa 7 µm), scarse in microfibrille e più mineralizzate. In queste ultime lamelle sono scavate le lacune ossee che accolgono i corpi degli osteociti. Questo modello consente anche di spiegare le notevoli proprietà di resistenza meccanica del tessuto osseo lamellare: le lamelle più ricche in microfibrille sarebbero infatti più plastiche e più adeguate ad assorbire urti e sollecitazioni meccaniche di tensione e di torsione, mentre quelle più ricche in minerale sarebbero più rigide e più adatte a sopportare senza deformarsi le forze di pressione applicate sul segmento osseo. Sovrapponendosi a strati, i due tipi di lamelle unirebbero le loro proprietà meccaniche realizzando un edificio leggero e molto resistente.
Gli osteoni appaiono come delle lunghe strutture cilindriche, il cui asse longitudinale tende a disporsi parallelamente alle linee di forza applicate su un dato segmento osseo. Così il femore, che sostiene il peso del corpo ed è quindi soggetto a forze applicate in senso longitudinale, è formato in prevalenza da osteomi, costituiti da microfibrille collagene, disposti parallelamente al suo asse maggiore.
Bibliografia:
· http://www3.unifi.it/anatistol/istologia/tessuto_osseo.htm, 21.04.2008
· http://www3.unifi.it/anatistol/istologia/figure_osseo.htm, 21.04.2008
· http://www.geol.lsu.edu/henry/Geology7900/Apatite/Apatite.htm, 04.05.2008
· http://www.nanopicoftheday.org/images/BoneApatite.jpg, 02.05.2008
· http://www.pforster.ch/UniTrieste/AnatomiaMartini/5/AM5_3.png, 10.05.2008
· http://webmineral.com/jpowd/JPX/jpowd.php?target_file=Carbonate-fluorapatite_2.jpx,
02.05.2008
· Alexander Spence, Basic Medical Anatomy, Benjamin/Cummings, 1990
· http://web.mit.edu/3.082/www/team1_f02/collagen.htm, 01.05.2008
Fonte: http://zyxel-nsa210.lilu2.ch/MyWeb/public/chimica/Devittori/LAM/LM08/nanotech/tessuto_osseo_CM.doc
Autore del testo: non indicato nel documento di origine
Le ossa
Nel mondo animale ci sono gli invertebrati e i vertebrati: noi esseri umani siamo vertebrati derivati dalle scimmie (primati) e contraddistinti per la posizione erettile che assumiamo grazie al nostro scheletro; nel nostro organismo lo scheletro e muscoli formano insieme il sistema locomotore.
Il tessuto osseo entra a far parte di tutte le ossa del nostro organismo: è un tessuto connettivo formato da una matrice ossea chiamata osseina, che è una sostanza proteica molto dura perché impregnata di sali minerali e soprattutto di sali di calcio, e da fibre di collagene che rendono questo tessuto elastico (tanto più elastico quando l’individuo è più giovane e tanto più rigido quando l’individuo è più anziano). Nella matrice ci sono tre tipi di cellule (deputate alla sua formazione): gli osteociti (cellule mature), osteoblasti (cellule neonate) e gli osteoclasti (cellule che modellano l’osso per esempio quando vi è la rottura e c’è la produzione di tessuto osseo che deve essere modellato). Il tessuto osseo si divide in tessuto osseo compatto (toc) e tessuto osseo spugnoso (tos).
Ultrastruttura (visibile al microscopio) di questi tessuti:
Il tessuto osseo compatto: questo tessuto rende l’osso compatto; la sua unità funzionale è l’osteone formato da una serie di lamelle concentriche di osseina che formano una struttura cilindrica all’interno della quale vi è il canale di Havers in cui scorrono vasi sanguigni a funzione nutritiva (funzione trofica) e terminazioni nervose che conferiscono all’osso la sensibilità. Gli osteoni comunicano per mezzo di altri canali.
Il tessuto osseo spugnoso: questo tessuto ha l’aspetto di una spugna perché è formato da bacchette di osseina che, intersecandosi tra loro, delimitano delle lacune al cui interno vi è il midollo osseo rosso con funzione emopoietica. Questo tessuto si trova per esempio nelle epifisi e nello sterno.
Oltre al tessuto osseo nel nostro organismo vi è un altro tessuto sempre connettivo chiamato cartilagine strettamente connessa al nostro scheletro: è formata dalla matrice (condrina), fibre di collagene e cellule. Alla nascita una larga parte delle nostre ossa sono formate da questo tessuto che successivamente si trasforma in osso con il processo di ossificazione. La cartilagine riveste le articolazioni (l’unione tra due ossa che possono essere fisse, mobili o meno mobili) ed ha la funzione di evitare l’attrito tra le ossa. La componente cellulare della cartilagine è rappresentata da tre tipi di cellule (deputate alla sua produzione): condrociti (cellule mature), condroblasti (cellule neonate o immature), condroclasti (cellule che distruggono la cartilagine durante i processi di ossificazione indiretta).
Ossificazione diretta e indiretta: l’ossificazione diretta avviene tramite produzione di osso direttamente sulla membrana, quella indiretta si attua per sostituzione della cartilagine con l’osso
Suddivisione generale delle ossa:
Le ossa si dividono in ossa lunghe, ossa brevi e ossa piatte.
Le ossa lunghe sono quelle degli arti: sono formate da una parte centrale tubolare chiamata diafisi, formata da tessuto osseo compatto al cui interno vi è un canale dove si trova il midollo giallo ricco di lipidi (funzione trofica), e due rigonfiamenti alle estremità chiamati epifisi . Finché l’individuo non ha completato la crescita tra le epifisi e la diafisi vi è una placca di cartilagine di coniugazione che serve per l’accrescimento dell’osso: parte di questa cartilagine viene ossificata tramite ossificazione indiretta (l’osso si forma dalla cartilagine e non direttamente), se ne forma poi dell’altra, si ossifica un’altra parte e così via durante tutto l’accrescimento che è controllato da un ormone somatotropo dell’adenoipofisi (GH). L’ipofisi è la ghiandola più importante del nostro organismo e si trova in posizione ipotalamica all’interno nella scatola cranica: è formata da una parte anteriore chiamata adenoipofisi (che appunto produce l’ormone) e una parte posteriore chiamata neuroipofisi. Questa ghiandola è situata nella sella turcica dello sfenoide, un osso a forma di farfalla con le ali e il cui corpo ha la forma di una sella che si trova alla base degli emisferi cerebrali.
Le ossa brevi sono per esempio le vertebre e si differenziano dalle altre per avere le tre dimensioni uguali.
Le ossa piatte sono ossa in cui una dimensione è molto più piccola delle altre due: sono formate di solito da due strati di tessuto osseo compatto e all’interno vi è il tessuto osseo spugnoso.
Alcune ossa del nostro scheletro:
La colonna vertebrale è formata nella maggior parte dei casi da 33 o 34 vertebre che si dividono per zona: cervicale (7), toracica (12), lombare (5), coccigea(5) e sacrale(4 o 5). La vertebra è costituita da un corpo vertebrale che insieme all’arco vertebrale delimita il foro vertebrale, il quale, insieme agli altri fori vertebrali, per sovrapposizione delle vertebre, costituisce il canale vertebrale all’interno del quale è contenuto il midollo spinale. Tra una vertebra e l’altra ci sono dei dischi fibrosi intervertebrali che attutiscono i movimenti: quando si fa uno sforzo, un movimento strano o si ha una postura sbagliata uno di questi dischi può essere schiacciato provocando l’ernia al disco.
Le ossa del cranio sono formate dalle ossa della faccia e della testa; le ossa della testa sono: l’osso frontale e l’occipitale posteriore (due ossa impari). L’occipitale è provvisto di un foro chiamato foramen magnum, attraverso il quale passano le strutture nervoso. Superiormente ci sono le parietali, inferiormente le temporali provviste di una struttura piramidale chiamata rocca petrosa che protegge parte dell’orecchio. Queste ossa sono legate tra loro mediante articolazioni fisse o chiamate suture. Le ossa della faccia sono invece: la mascella, le ossa zigomatiche,la mandibola, le ossa nasali, le ossa lacrimali ecc. ecc.
La gabbia toracica protegge alcuni organi del nostro organismo come i polmoni e il cuore: è formata da dodici paia di costole o coste che posteriormente sono articolate con le faccette articolari delle vertebre mentre anteriormente allo sterno. Le prime 7 sono vere perché si articolano direttamente allo sterno poi ne abbiamo 3 false perché unite alla cartilagine costale delle settime costole e due paia sono fluttuanti ovvero non sono articolate anteriormente per permettere una più ampia espansione della respirazione. Lo sterno consiste in tre parti: il manubrio, il corpo e il processo xifoideo.
Il cinto scapolare e arto superiore: è rappresentato dalla spalla che è formata anteriormente dalla clavicola (un osso lungo) e posteriormente dalla scapola, al cinto scapolare si articola l’omero che è l’osso del braccio mentre le ossa dell’avambraccio si chiamano ulna e radio, quelle della mano sono il carpo e il metacarpo quelle delle dita sono la falange, la falangina e la falangetta (tutte ossa lunghe).
Il cinto pelvico e arto inferiore è formato dalle ossa dell’anca: l’ileo, un grosso osso che ha la forma piatta, l’ischio e il pube che si fonde con il pube simmetrico in una sutura chiamata sinfisi pubica. L’anca si articola con la testa del femore che è l’osso della coscia tramite la fossa dell’acetabolo, cavità glenoidea formata dalla fusione di ileo, ischio e pube, inferiormente al femore troviamo il ginocchio e le ossa della gamba rappresentate dalla tibia e dal perone, a queste seguono le ossa del piede che sono il tarso e il metatarso e quelle delle dita del piede che sono la falange, la falangina e la falangetta. L’alluce a differenza del pollice ha solo la falange e la falangina.
Fonte: http://classi3webios.myblog.it/media/00/01/763099247.doc
Autore del testo: non indicato nel documento di origine
Tessuto osseo
Visita la nostra pagina principale
Tessuto osseo
Termini d' uso e privacy