Biologia appunti parte 1
Biologia appunti parte 1
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
BIOLOGIA
Tutti gli esseri viventi sono costituiti da una (unicellulari) o più (pluricellulari) cellule.
La cellula può essere considerata come la più piccola porzione di un essere vivente in grado di conservare le caratteristiche della vita (nutrizione, riproduzione etc.).
La citologia è la branca della biologia che studia la cellula.
Le cellule vennero osservate per la prima volta nel 1600 da Robert Hooke, il quale, analizzando attraverso un microscopio (strumento da poco inventato) sottili sezioni di sughero, mise in evidenza l'esistenza di piccole cellette, che egli chiamò appunto cellule (dal latino “cellula” = piccola cella).
Ma quella che oggi chiamiamo "teoria cellulare" si affermò solo nella prima metà dell'Ottocento ad opera di Schleiden, Schwann e Virchow. Nella sua formulazione moderna la teoria cellulare afferma che tutti gli esseri viventi sono costituiti da cellule e che ogni cellula si genera da altre cellule.
La cellula, come tutta la materia ordinaria, é costituita di sostanze chimiche.
Lo studio dei composti chimici e dei fenomeni fisico-chimici che stanno alla base della vita ha assunto un'importanza via via maggiore, tanto che oggi vi é addirittura una branca della chimica moderna che se ne occupa, la biochimica.
Inoltre alcuni composti chimici quali ad esempio gli acidi nucleici (RNA e DNA) e le proteine, vengono sintetizzati solo dagli esseri viventi e sono talmente importanti nel regolare tutte le loro attività da essere studiati dalla biologia molecolare.
La biochimica e la biologia molecolare rappresentano un tentativo riduzionistico ed in qualche modo meccanicistico di spiegare il tutto attraverso l'analisi delle sue parti elementari e dei modi in cui queste si integrano e si organizzano.
Il livello di organizzazione cellulare manifesta in altre parole un livello di organizzazione sottostante che potremo definire livello molecolare. E questo a sua volta un livello inferiore detto livello atomico.
La materia si struttura organizzandosi attraverso più livelli a complessità crescente e la scienza moderna cerca di spiegare ciascun livello, "riducendolo" a quello più elementare che lo precede, in un gioco delle scatole cinesi che non ha finora ancora trovato la sua fine.
Così il livello cellulare viene "ridotto" al livello molecolare. Quello molecolare viene interpretato in base al sottostante livello atomico, per giungere a quello che per ora sembra essere il livello più semplice e che chiameremo genericamente livello subatomico.
Anche se ogni livello è formato dall'ordinata aggregazione ed organizzazione delle strutture che costituiscono il livello sottostante, esso non può mai essere descritto come semplice somma degli elementi più semplici che lo formano. Ciascun livello è infatti caratterizzato da nuove proprietà non descrivibili e soprattutto non prevedibili sulla base delle semplici caratteristiche degli elementi che lo compongono (proprietà emergenti).
Così la molecola d'acqua (livello molecolare) presenta caratteristiche totalmente nuove e diverse rispetto a quelle degli atomi di Idrogeno e di Ossigeno (livello atomico) che la compongono.
Ma forse la caratteristica più spettacolare che compare a livello cellulare e che non è nemmeno lontanamente immaginabile studiando il sottostante livello molecolare è ciò che noi chiamiamo vita.
E' quindi probabilmente illusorio sperare di spiegare che cos'è la vita studiando le molecole, i composti chimici di cui è fatta una cellula. Ce lo conferma, se ce ne fosse bisogno, il fatto che, dopo aver smontato una cellula nei composti chimici che la costituiscono, non siamo più in grado di ricostruirla.
Nonostante ciò l'approccio riduzionistico e analitico ci ha comunque permesso di allargare enormemente le nostre conoscenze sui meccanismi e le strutture che stanno alla base di un essere vivente. Tanto che oggi é impossibile iniziare lo studio della biologia in generale e della citologia in particolare senza affrontare preliminarmente gli elementi fondamentale della chimica della vita.
Elementi di biochimica
Le innumerevoli sostanze chimiche che costituiscono tutti gli oggetti, naturali e artificiali che compongono l'universo sono a loro volta costituite dalla combinazione di 92 tipi di atomi diversi.
Il termine atomo significa indivisibile e fu coniato dal filosofo greco Democrito per indicare l'esistenza in natura di particelle elementari, non ulteriormente divisibili.
In realtà i 92 tipi di materia che costituiscono l'universo non sono atomi in senso etimologico poiché, come vedremo, possono essere ulteriormente spezzati in particelle più piccole.
Il termine é comunque entrato stabilmente nella terminologia scientifica.
Struttura atomica
Tutti gli atomi sono formati da 3 tipi di particelle più piccole (particelle subatomiche):
- Il protone il quale possiede la più piccola quantità di carica positiva finora osservata in natura (1,6.10-19 coulomb) e massa pari a 1,67.10-24 g.
- Il neutrone il quale non possiede carica elettrica e presenta una massa dello stesso ordine di grandezza del protone.
- l'elettrone il quale possiede la più piccola quantità di carica elettrica negativa finora osservata, pari ma di segno opposto a quella del protone. L'elettrone presenta una massa circa 2000 volte inferiore rispetto a quella di un protone (1/1836).
Ricordiamo che cariche dello stesso segno si respingono, mentre cariche di segno contrario si attraggono.
Gli atomi sono costituiti da una massiccia parte centrale positiva, formata da protoni e neutroni, detta nucleo, e da una nuvola di elettroni che orbitano a grandi distanze dal nucleo a velocità enormi dell'ordine di 108 km/s.

Si noti come non sempre la misura precisa di una grandezza fisica sia interessante. A volte è sufficiente conoscere il suo ordine di grandezza. In tal caso è necessario esprimere le sue dimensioni come una potenza di 10 (notazione esponenziale). Il confronto tra ordini di grandezza è più significativo ed immediato, essendo sufficiente eseguire la differenza tra gli esponenti del 10.
Ad esempio, se vogliamo avere un’idea delle proporzioni di un atomo, possiamo confrontare le sue dimensioni con quelle del suo nucleo. Dunque praticamente tutta la massa dell'atomo é concentrata nel suo nucleo, in una regione delle dimensioni dell’ordine di 10-13 cm.
Ma il volume dell'atomo é costituito dalle orbite elettroniche, le quali danno all'atomo una dimensione totale dell'ordine di 10-8 cm (1 ångstrom), 5 ordini di grandezza superiore a quelle del nucleo (13-8=5).
Per renderci conto della distanza esistente tra nucleo ed elettroni, basti pensare che se il nucleo avesse un diametro di 1 cm, gli elettroni dovrebbero girare ad una distanza 105 = 100.000 volte maggiore, pari a 100.000 cm = 1 km. In tal modo possiamo affermare che la materia comune é formata più di vuoti che di pieni, una specie di colabrodo la cui apparenza di solidità é legata al moto vorticoso degli elettroni (un po' come un'elica di un aereo che, girando, ci appare come un muro impenetrabile).
Normalmente un atomo possiede tanti elettroni quanti protoni, per cui, complessivamente, esso risulta neutro.
Gli atomi si legano tra loro a formare i diversi composti chimici rimanendo a contatto con la superficie del guscio elettronico (i gusci elettronici non permettono infatti agli atomi di avvicinarsi oltre una certa distanza in virtù della repulsione elettrostatica esistente tra cariche dello stesso segno).
Cosa distingue dunque i 92 tipi di atomi tra loro, dato che sono costituiti tutti di protoni, neutroni ed elettroni? Cosa rende un atomo di Ferro diverso da un atomo di Oro?
La risposta é il numero di protoni. Tale numero é una specie di carta di identità per l'atomo e viene chiamato numero atomico Z (dal tedesco Zahl = numero).
Un atomo con 6 protoni é solo ed esclusivamente l'atomo di Carbonio (simbolo chimico C).
Un atomo con 8 protoni é solo l'atomo di Ossigeno (simbolo chimico O).
Elementi e composti
I 92 atomi possiedono un numero di protoni che va da 1 (per l'Idrogeno H) fino a 92 (per l'Uranio U). I 92 tipi di atomi corrispondono ad altrettanti elementi diversi. Il termine “elemento” indica una sostanza materiale “non composta” da sostanze più semplici
Così l’acqua non è un elemento, ma un composto, essendo costituita da due sostanze semplici o elementari: l’idrogeno (2 atomi) e l’ossigeno (1 atomo).
Il numero atomico (Z) viene posto in basso a sinistra del simbolo dell'elemento. Ad esempio 6C indica che il carbonio ha numero atomico 6 e contiene quindi 6 protoni nel nucleo.
In modo analogo, con un numero posto in alto a sinistra del simbolo dell’elemento si indica il numero di massa (A), che è dato dalla somma del numero di protoni (Z) e di neutroni (N) contenuti nel nucleo. Il simbolo 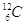 (carbonio sei-dodici) ci informa che l’atomo di carbonio possiede 6 protoni e 6 neutroni (N = A - Z = 12 - 6).
(carbonio sei-dodici) ci informa che l’atomo di carbonio possiede 6 protoni e 6 neutroni (N = A - Z = 12 - 6).
Isotopi e decadimenti
Gli elementi che hanno lo stesso numero atomico, ma diverso numero di massa si dicono isotopi. Un particolare isotopo di un elemento si indica in genere utilizzando solo il numero di massa, dando per sottinteso il numero atomico. Così 14C (carbonio quattordici) indica l'isotopo del carbonio che contiene 8 neutroni, anziché i 6 dell'isotopo più stabile, che è appunto il 12C (carbonio dodici).
Negli elementi con numero atomico piccolo (Z < 20), il numero dei neutroni nell'isotopo più stabile è generalmente uguale a quello dei protoni o vi differisce per una unità in eccesso. Al crescere di Z, il rapporto fra N e Z aumenta progressivamente. È probabile che i neutroni abbiano un ruolo importante nel dare stabilità ai nuclei. Mentre ad esempio il Carbonio possiede 6 p e 6 n, l'Ossigeno 8 p e 8 n, il Ferro presenta 26 p e 30 n, mentre l'Uranio 92 p e 146 n.
I nuclei degli elementi più pesanti sono in genere instabili e tendono a decadere emettendo particelle per raggiungere una condizione di maggior stabilità nucleare. È stato osservato che i nuclei con valori pari di Z e N sono più stabili di quelli con valori dispari.
La maggior parte degli elementi è presente in natura sottoforma di miscele di isotopi.
Ad esempio il gas Idrogeno H2 è costituito da una miscela di 3 isotopi
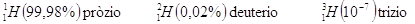
Alcuni isotopi sono instabili, sono cioè soggetti a decadimento radioattivo (radioisotopi). Il decadimento radioattivo comporta l'emissione di particelle energetiche con trasformazione degli isotopi instabili in isotopi di elementi diversi.
Vi sono due tipi fondamentali di decadimenti: alfa e beta.
1) decadimento b (beta): un neutrone del nucleo si trasforma in un protone (che rimane nel nucleo, aumentando di un'unità il numero atomico e quindi trasformando l'elemento in quello che lo segue nella tabella periodica), e in un elettrone ed un antineutrino (che si allontanano dal nucleo ad elevata velocità).
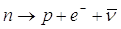
Nel decadimento b inverso un protone del nucleo, colpito da un elettrone, si trasforma in un neutrone (che rimane nel nucleo, diminuendo di un'unità il numero atomico e quindi trasformando l'elemento in quello che lo precede nella tabella periodica) e in un neutrino (che si allontana dal nucleo).
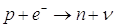
Una reazione equivalente prevede che un protone emetta un positrone (l'antiparticella dell'elettrone) ed un neutrino trasformandosi in un neutrone, che rimane nel nucleo.
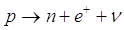
2) decadimento a (alfa): il nucleo di un isotopo espelle un nucleo di Elio (2 protoni + 2 neutroni) o particella a (diminuendo di due unità il proprio numero atomico e trasformandosi nell'elemento che lo precede di due posti nella tabella periodica).
Molti radioisotopi hanno un'interesse rilevante nella ricerca scientifica.
Un'applicazione notevole si ha nella datazione di rocce e fossili. Il metodo si basa sull'osservazione che ciascun radioisotopo impiega un tempo ben determinato per trasformarsi, in genere attraverso una lunga serie di elementi intermedi instabili, in un isotopo stabile. Tutti gli isotopi radioattivi decadono obbedendo alla stessa legge di decadimento:
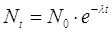
dove
N0 = numero di atomi iniziali
Nt = numero di atomi residui (che non hanno ancora subito il decadimento) dopo un tempo t
l = costante di decadimento (diversa da elemento ad elemento)
t = tempo
Ponendo 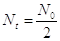 ) o di semitrasformazione o emivita, cioè il tempo necessario affinchè decadano metà degli atomi iniziali. La legge di decadimento diventa
) o di semitrasformazione o emivita, cioè il tempo necessario affinchè decadano metà degli atomi iniziali. La legge di decadimento diventa
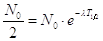
ed il tempo di dimezzamento

I fisici hanno determinato con grande accuratezza il tempo di dimezzamento dei radioisotopi.
isotopo instabile |
decadimento |
isotopo stabile |
tempo di dimezz. |
|
8a e 6b |
|
4,51 miliardi |
|
7a e 4b |
|
713 milioni |
|
6a e 4b |
|
13,9 miliardi |
|
b inverso |
|
1,26 miliardi |
|
b |
|
50 miliardi |
|
b |
|
5.730 |
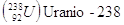
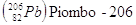
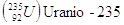
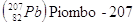

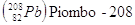


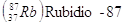
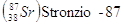

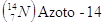
L'attività di una sostanza radioattiva si misura in curie.
1 curie = 3,7.1010 decadimenti a o b al secondo.
Misurando dunque la percentuale di un certo isotopo e quella dei suoi prodotti di decadimento presenti in una roccia è possibile risalire alla sua età.
Se infatti chiamiamo G la quantità dell'isotopo genitore residuo e F la quantità dell'isotopo figlio che si è formato, la legge di decadimento può essere scritta
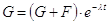
e poiché la costane di decadimento vale
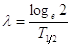
sostituendo, otteniamo
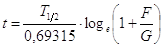
E' evidente dunque che per conoscere l'età t di una roccia è sufficiente conoscere il tempo di dimezzamento di un certo radioisotopo e misurare il rapporto isotopico tra elemento figlio ed elemento genitore che rappresenta una specie di memoria della storia chimica della roccia.
Per datare rocce molto antiche si utilizza in genere il metodo Rubidio-Stronzio, mentre per rocce aventi un'età compresa tra 200.000 e 1 miliardo di anni è preferibile il metodo Potassio-Argon.
Se ad esempio in una roccia misuriamo un rapporto isotopico 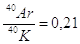 , l'età della roccia potrà essere calcolata in
, l'età della roccia potrà essere calcolata in
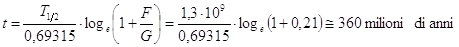
Per la datazione di resti organici si utilizza il metodo del Carbonio-14.
Il carbonio 14 si produce continuamente nell'alta atmosfera per interazione dei gas atmosferici con i raggi cosmici. Da tali interazioni si producono neutroni liberi (per disintegrazione di nuclei gassosi) che possono al loro volta reagire con nuclei di azoto atmosferico, secondo la reazione
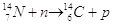
Il Carbonio-14, una volta formatosi, decade nuovamente in Azoto-14 (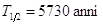 ), emettendo un elettrone ed un antineutrino
), emettendo un elettrone ed un antineutrino
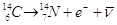
La velocità delle due reazioni è tale per cui la quantità di Carbonio-14 presente nell'atmosfera rimane costante ed il rapporto Carbonio-12/Carbonio-14 presenta un valore ben definito, pari a
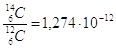
circa 1 atomo di C-14 ogni 1000 miliardi di atomi di C-12.
Il valore di tale rapporto si mantiene inalterato anche durante i processi di fissazione fotosintetica della CO2 da parte dei vegetali ed in generale di assimilazione da parte degli organismi che direttamente o indirettamente si nutrono di vegetali. In altre parole finché un qualsiasi organismo animale o vegetale è vivo esso risulta in equilibrio con l'ambiente e quindi al suo interno il rapporto 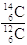 si mantiene costante. Dal momento in cui un organismo muore l'assunzione di C-14 cessa, mentre il suo decadimento in N-14 prosegue.
si mantiene costante. Dal momento in cui un organismo muore l'assunzione di C-14 cessa, mentre il suo decadimento in N-14 prosegue.
E' allora evidente che la misurazione del rapporto 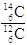 all'interno di un reperto organico ci permette di risalire alla data della sua morte.
all'interno di un reperto organico ci permette di risalire alla data della sua morte.
Tenuto comunque conto della relativa brevità del tempo di dimezzamento del C-14, tale metodo non permette di datare fossili più vecchi di circa 50-70.000 anni.
In campo medico e biologico molte vie metaboliche sono state scoperte utilizzando composti "marcati" con radioisotopi, seguendone poi il destino attraverso complicate sequenze di reazioni all'interno dell'organismo. Se infatti mettiamo a disposizione dell'organismo isotopi radioattivi dell'Ossigeno, dell'Azoto o del Carbonio, questi, possedendo le medesime caratteristiche chimiche dei rispettivi isotopi stabili, vengono normalmente assimilati ed utilizzati nella sintesi delle sostanze necessarie. Tali sostanze risultano a questo punto radioattive e quindi facilmente localizzabili ed isolabili.
Sempre in campo medico le radiazioni emesse dalla disintegrazione del Cobalto-60 (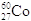 ) essendo chimicamente affine al Calcio (appartiene allo stesso gruppo chimico) può essere utilizzato dall'organismo al posto di quest'ultimo all'interno del tessuto osseo, provocando l'insorgenza di forme tumorali, specialmente negli organismi in fase di crescita. Inoltre il suo tempo di dimezzamento è di circa 28 anni, sufficientemente lungo per concentrarsi pericolosamente nell'ambiente ed essere assorbito dagli esseri viventi.
) essendo chimicamente affine al Calcio (appartiene allo stesso gruppo chimico) può essere utilizzato dall'organismo al posto di quest'ultimo all'interno del tessuto osseo, provocando l'insorgenza di forme tumorali, specialmente negli organismi in fase di crescita. Inoltre il suo tempo di dimezzamento è di circa 28 anni, sufficientemente lungo per concentrarsi pericolosamente nell'ambiente ed essere assorbito dagli esseri viventi.
La tabella periodica
I 92 tipi di atomi o elementi sono stati ordinati in una tabella in ordine di numero atomico crescente, dal più leggero, l'Idrogeno, al più pesante, l'Uranio (il peso atomico dipende essenzialmente dal numero totale dei protoni e dei neutroni (nucleoni) che si trovano nel nucleo e quindi dal numero atomico).
La tabella é stata impostata verso la metà dell'Ottocento (1869) dal chimico russo Mendeleev (il quale inizialmente ordinò gli elementi secondo il peso atomico crescente).
Mendeleev si accorse che, passando da elementi più leggeri ad elementi a più elevato peso atomico, le caratteristiche chimiche già incontrate in precedenza si ripresentavano periodicamente negli elementi più pesanti.
Mendeleev sistemò allora gli elementi in una tabella formata da un certo numero di righe e di colonne, in modo tale che gli elementi che presentavano proprietà simili si incolonnassero.
Spostandosi lungo una linea orizzontale (periodo) il numero atomico cresce di una unità per volta e le proprietà chimiche variano con continuità.
Scendendo lungo una colonna (gruppo) il numero atomico cresce di parecchie unità per volta, ma le proprietà chimiche rimangono praticamente costanti.
E' un po' come sistemare un mazzo di carte mettendo tutte le carte di uno stesso seme, dalla più piccola alla più grande, su quattro righe orizzontali, in modo tale che gli assi si incolonnino con gli assi, i due con i due e così via.

La somiglianza chimica tra elementi appartenenti ad un medesimo gruppo é dovuta al tipo di distribuzione degli elettroni attorno al nucleo.
Livelli energetici ed orbitali
Gli elettroni orbitano intorno al nucleo secondo schemi particolari, scoperti solamente nella prima metà del Novecento. Inizialmente si riteneva che gli elettroni percorressero traiettorie definite all’interno di orbite circolari ed ellittiche. Successivamente si è scoperto che è impossibile definire con precisione la traiettoria di un elettrone ed il concetto di orbita è stato sostituito dal concetto di orbitale. L’orbitale è la regione di spazio intorno al nucleo alla quale si associa una certa probabilità, arbitrariamente fissata (in genere del 90-95%) di trovare l’elettrone. Gli orbitali vengono rappresentati tramite gusci delimitati da superfici chiuse o tramite nuvole sfumate (nuvola elettronica) la cui intensità di colore è proporzionale alla probabilità di trovare l’elettrone. Vi sono 4 tipi di orbitali di forma diversa, indicati con le lettere s, p, d, f. Gli orbitali s sono sferici, gli orbitali p sono bilobati, mentre gli orbitali d ed f hanno complesse forme multilobate.
Ogni orbitale può contenere al massimo due elettroni.

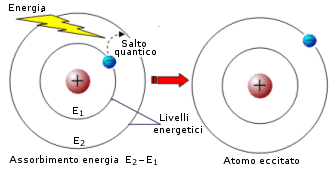
Gli elettroni possono sistemarsi solo a certe distanze dal nucleo, formando così una sorta di strati o gusci elettronici, a ciascuno dei quali compete una certa energia caratteristica, detta livello energetico. I livelli energetici sono quantizzati, possono cioè possedere solo certi valori di energia. Ogni livello energetico può contenere solo un certo numero di elettroni, caratteristico del livello. Gli elettroni tendono ad occupare sempre il livello energetico libero più basso (più vicino al nucleo) fino a saturarlo e solo allora si dispongono in livelli energetici più esterni.
Un elettrone può passare da un livello più interno di energia Ei ad uno più esterno libero di energia Ee, solo se gli viene fornita esattamente la quantità di energia Ee-Ei. In altre parole se sono quantizzati i livelli energetici, allora sono quantizzate anche le differenze di energia tra i livelli (dislivelli energetici quantizzati). Ciò significa che ogni atomo può assorbire solo certe quantità di energia caratteristiche. Come abbiamo appena detto, quando un atomo assorbe energia un elettrone passa da un livello più interno ad uno più esterno (salto quantico o transizione elettronica) e l’atomo si viene a trovare in una situazione di instabilità (stato eccitato). Quando l’elettrone ritorna al livello energetico di partenza, viene restituita esattamente la medesima quantità di energia assorbita e l’atomo riacquista stabilità (stato fondamentale).
Nel primo livello energetico, quello più vicino al nucleo, possono essere ospitati non più di 2 elettroni (vi è solo un orbitale s)
Nel secondo livello energetico altri 2 elettroni come in precedenza più altri 6. Si tratta in realtà di due sottolivelli il primo con 2 elettroni (1 orbitale s) ed il secondo con 6 elettroni (3 orbitali p) per un totale di 8 elettroni.
Nel terzo livello 2 + 6 + 10 elettroni (3 sottolivelli: 1 orbitale s + 3 orbitali p + 5 orbitali d) per un totale di 18 elettroni.
Nel quarto livello 2 + 6 + 10 + 14 elettroni (4 sottolivelli: 1 orbitale s + 3 orbitali p + 5 orbitali d + 7 orbitali f) per un totale di 32 elettroni.
I livelli successivi presentano al massimo 4 sottolivelli. Gli atomi più pesanti, come l’Uranio, hanno elettroni a sufficienza per occupare 7 livelli energetici, senza tuttavia riuscire a riempirli completamente.
Gli atomi utilizzano gli elettroni del loro livello energetico più esterno (elettroni superficiali o elettroni di valenza) per interagire tra loro. Il tipo di reazioni che possono fare dipende quindi dal numero di elettroni del loro strato di valenza o, come si usa dire, dalla loro configurazione elettronica superficiale. Atomi di elementi diversi che presentino la medesima configurazione elettronica superficiale (il medesimo numero di elettroni sul loro ultimo livello) manifestano quindi caratteristiche chimiche simili.
Così l'Idrogeno, il primo e più leggero degli elementi chimici é costituito di 1 protone nel nucleo e di un elettrone nel primo livello energetico.
L'Elio con 2 protoni + 2 neutroni nel nucleo e due elettroni orbitanti, satura il primo livello energetico.
Il Litio, che possiede 3 protoni, 3 neutroni e tre elettroni, sistema i primi due elettroni nel primo livello, saturandolo, ed é costretto a sistemare il suo terzo elettrone nel secondo livello. per questo motivo viene incolonnato sotto l'Idrogeno. Come l'idrogeno presenta infatti un unico elettrone sul livello energetico più superficiale.
L'elemento successivo, il Berillio (Be 4 elettroni), ne sistema 2 nel primo livello e 2 nel secondo.
Il Boro (B - 5 elettroni) ne sistema 2 nel primo e 3 nel secondo.
Il Carbonio (C - 6 elettroni) ne sistema 2 nel primo e 4 nel secondo.
L'Azoto (N - 7 elettroni) 2 nel primo e 5 nel secondo.
L'Ossigeno (O - 8 elettroni) 2 nel primo e 6 nel secondo.
il Fluoro (F - 9 elettroni) 2 nel primo e 7 nel secondo.
Il Neon (Ne - 10 elettroni), con 2 elettroni nel primo e 8 nel secondo, satura il secondo livello energetico. Quando un elemento chimico satura con 8 elettroni i primi due sottolivelli del suo ultimo livello risulta chimicamente stabile. In altre parole non manifesta alcuna tendenza a modificare la sua configurazione elettronica, tramite reazioni chimiche con altri atomi. Il Neon si trova in questa condizione.
Il Sodio (Na), che possiede 11 elettroni ne sistema 2 nel primo, 8 nel secondo e 1 nel terzo.
Il Sodio viene a possedere così un unico elettrone sul livello energetico più superficiale, come il Litio e l'Idrogeno e viene perciò incolonnato sotto di essi.
Il Magnesio (Mg) che possiede 12 elettroni ne sistema 2 nel primo livello, 8 nel secondo e 2 nel terzo. Possedendo 2 elettroni sul livello energetico più esterno, come il Berillio viene incolonnato sotto di esso.
Sotto il Magnesio, incolonnato nel suo stesso gruppo, troviamo il Calcio che dopo aver saturato di elettroni i primi tre livelli, presenta 2 elettroni nel quarto livello, quello più esterno e quindi presenta la stessa configurazione elettronica superficiale del Berillio e del Magnesio.
Tutti i successivi elementi si troveranno dunque incolonnati in modo tale da far parte di un gruppo di elementi i quali possiedono tutti uno stesso numero di elettroni sul livello energetico più superficiale. Naturalmente il livello energetico sarà diverso da elemento a elemento.
Dunque le caratteristiche simili degli elementi che appartengono ad uno stesso gruppo chimico dipendono essenzialmente dal numero di elettroni presenti sul livello energetico più superficiale, indipendentemente dal fatto che questo sia il primo, il secondo o l'ultimo.
Gli elementi del I gruppo A hanno tutti 1 elettrone superficiale
Gli elementi del II gruppo A hanno tutti 2 elettroni superficiali
Gli elementi del III gruppo A hanno tutti 3 elettroni superficiali
E così via fino agli elementi del gruppo 0 (zero) che, avendo 8 elettroni superficiali, sono tutti stabili (gas nobili)
Come abbiamo già detto, gli atomi reagiscono tra loro utilizzando gli elettroni più superficiali, mentre gli elettroni interni non partecipano in genere alle reazioni chimiche. Inoltre gli elementi che, come il Neon, possiedono 8 elettroni superficiali (1 orbitale s e 3 orbitali p saturi) risultano particolarmente stabili, inerti, nel senso che hanno pochissima tendenza a reagire con altri elementi chimici.
Gli altri elementi che possiedono configurazioni elettroniche simili a quella dei gas nobili tendono a perdere o ad acquistare elettroni per acquisire tale configurazione particolarmente stabile. Vedremo che molte reazioni chimiche possono essere spiegate proprio in virtù della tendenza di molti elementi ad acquisire la configurazione ad 8 elettroni superficiali dei gas nobili (regola dell'ottetto).
Simbologia chimica
Ogni elemento chimico viene convenzionalmente indicato con un simbolo chimico di una o due lettere, di cui la prima maiuscola.
Quando due o più atomi si uniscono si parla di molecola. Se una molecola è formata da atomi dello stesso elemento si parla di sostanza semplice o elementare.
Fe O2 H2 Au N2 S8
ferro ossigeno idrogeno oro azoto zolfo
L'indice posto in basso a destra indica il numero di atomi legati a formare una molecola. Quando non è presente è sottinteso l'indice 1.
Se una molecola è formata da atomi di elementi diversi si parla di sostanza composta o, più semplicemente, di composto.
H2O CO2 NH3 CH4 H3PO4
acqua anidride carbonica ammoniaca metano acido fosforico
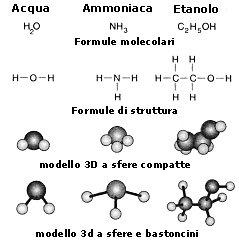
Le formule chimiche fin qui usate si dicono formule brute o grezze. Esse ci informano solo sul numero di atomi di ciascun elemento che entrano a far parte di una sostanza, ma non dicono nulla sul modo in cui gli atomi si legano tra loro.
Le formule che utilizzano delle barrette per evidenziare il legame chimico tra gli atomi sono dette formule di struttura. Esistono poi delle rappresentazioni tridimensionali (3D) che permettono di evidenziare la disposizione degli atomi nello spazio.
Quando si scrive una reazione chimica le sostanze che reagiscono (reagenti) vengono separate dalle sostanze che si formano (prodotti di reazione) dal segno di reazione (→).
Sia i reagenti che i prodotti di reazione sono preceduti da un numero, detto coefficiente stechiometrico, che indica il numero di molecole che partecipa alla reazione. Il coefficiente 1 è sottinteso.
2H2 + O2 → 2H2O
Due molecole di idrogeno (biatomico) reagiscono con una molecola di ossigeno (biatomico) per dare due molecole di acqua. Una reazione in cui compaiano i corretti coefficienti stechiometrici si dice “bilanciata”.
I coefficienti stechiometrici sono necessari al fine di soddisfare il principio di conservazione della massa. Durante la reazione infatti non si possono creare e non possono sparire atomi. Il numero di atomi di un elemento che si trova tra i reagenti deve sempre essere uguale al numero di atomi dello stesso elemento che si trova tra i prodotti di reazione.
Una reazione bilanciata è corretta non solo dal punto di vista qualitativo, ma anche da quello quantitativo e ci fornisce dunque informazioni sui rapporti con con cui le sostanze reagiscono.
Il peso (massa) degli atomi e delle molecole
Il numero di massa A non deve essere confuso con la massa effettiva di un elemento, in quanto, come abbiamo visto, ogni elemento è costituito da miscele di isotopi e quindi la sua è la massa media degli isotopi che lo compongono. Per indicare le masse degli atomi (o dei composti chimici) sarebbe scomodo usare l'unità di misura ordinaria della massa, il kg o il g. Per questo motivo si utilizza una unità di misura relativa, prendendo come riferimento la massa del 12C.
Il dalton o unità di massa atomica (uma o u) viene definito come 1/12 (un dodicesimo) della massa del C-12 ed è pari a 1,6605.10-24 g.
Si definisce peso atomico (o molecolare) relativo il rapporto tra la massa di un atomo (o di una molecola) e 1/12 della massa del Carbonio-12. Il peso risulta quindi essere "relativo" alla massa del C-12, presa arbitrariamente e convenzionalmente come unità di misura.
Così quando affermiamo che l'Ossigeno ha un peso atomico relativo di 16 dalton (o uma o u) ciò significa che esso pesa 16 volte più di 1/12 del C-12.
Il termine "peso" andrebbe più correttamente sostituito con il termine "massa", ma è ormai entrato nell'usuale terminologia chimica.
I pesi atomici relativi sono tabulati nella tabella periodica. Per calcolare i pesi molecolari relativi è sufficiente sommare i pesi atomici relativi di tutti gli atomi presenti in una molecola.
Ad esempio sapendo che l'Ossigeno pesa 16 u e l'Idrogeno 1 u, una molecola d'acqua H2O peserà 18 u (1 + 1 + 16), mentre una molecola di anidride carbonica CO2 peserà 44 u (12 + 16 + 16).
Un’altra unità di misura, usata per esprimere quantità macroscopiche di materia, è la mole.
1 mole di una sostanza è pari al suo peso relativo espresso in grammi.
Così una mole di ossigeno gassoso O2 è pari a 32 g di ossigeno (il peso relativo è 16 + 16 = 32 u). 1 mole di acqua è pari a 18 g di acqua, 1 mole di anidride cabonica è pari a 44 g di anidride carbonica.
Una mole di una qualsiasi sostanza ha la proprietà notevole di contenere sempre lo stesso numero di particelle, detto numero di Avogadro, pari a 6,022.1023. Così 32 g di ossigeno e 44 g di anidride carbonica contengono sempre un numero di Avogadro di molecole.
Tale proprietà è fondamentale nelle reazioni chimiche. Prendiamo ad esempio la reazione di sintesi dell'acqua dai suoi elementi
2H2 + O2 ð 2H2O
I coefficienti stechiometrici mi informano che l'idrogeno e l'ossigeno reagiscono nel rapporto numerico (molecolare) di 2:1. In altre parole, per ogni due molecole di idrogeno che reagiscono con una molecola di ossigeno si ottengono due molecole di acqua.
Ma potremo anche affermare che due moli di idrogeno (4 g) reagiscono con una mole di ossigeno (32 g) per dare 2 moli di acqua (36 g). E ciò perchè 2 moli di idrogeno contengono estattamente il doppio di molecole di 1 mole di ossigeno e quindi utilizzando le moli il rapporto molecolare di 2:1 viene comunque rispettato.
Il vantaggio sta nel fatto che ora possiamo misurare e far reagire quantità macroscopiche e facilmente misurabili di materia (4 g di idrogeno e 32 g di ossigeno).
Il legame chimico
La vita di una cellula dipende in larga misura dalla sua capacità di immagazzinare ed utilizzare energia. L'unico modo che la cellula possiede per immagazzinare energia è quella di fissarla all'interno dei legami chimici che tengono uniti gli atomi nelle molecole.
E' infatti attraverso una continua rottura e ricostruzione dei legami chimici e quindi attraverso un lavoro ininterrotto di demolizione e di riassemblaggio di molecole (metabolismo cellulare) che la cellula fa circolare l'energia al suo interno.
Una delle caratteristiche importanti di ogni legame chimico è dunque proprio il suo contenuto energetico, comunemente (anche se impropriamente) detto forza di legame.
Durante una reazione chimica vengono rotti alcuni legami dei reagenti, mentre si formano altri legami necessari per la sintesi dei prodotti di reazione.
Una reazione avviene con liberazione di energia (reazione esoergonica) se i legami che si spezzano sono più energetici dei legami che si formano. In tal caso possiamo affermare che i reagenti sono più energetici dei prodotti di reazione.
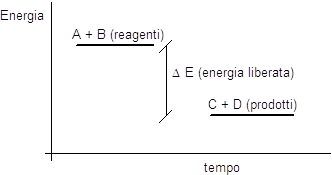
Se al contrario una reazione avviene con assorbimento di energia (reazione endoergonica) e i legami che si spezzano sono meno energetici dei legami che si formano, allora possiamo affermare che i reagenti sono meno energetici dei prodotti di reazione.

Le variazioni energetiche (DE) associate alla rottura ed alla formazione dei legami chimici si misurano normalmente in chilocalorie su mole (kcal/mol) o chilojoule su mole (kj/mol)
Ricordiamo che una caloria è la quantità di calore necessaria per far aumentare la temperatura di 1 g di acqua, portandola da 14,5°C a 15,5°C e che 1 cal = 4,186 J
Un esempio di reazione esoergonica è la combustione del metano in presenza di ossigeno.
CH4 + 2O2 ð CO2 + 2H2O + 212,8 kcal
Ogni mole di metano (16 g) che brucia libera dunque 212,8 Kcal
Un esempio di reazione endoergonica è la scissione dell'acqua in idrogeno ed ossigeno
2H2O + 136,6 Kcal ð 2H2 + O2
Per spezzare una mole di acqua (18 g) è necessario dunque fornire 68,3 Kcal.
Come si è già avuto modo di dire la reattività di un elemento chimico dipende essenzialmente dalla sua configurazione elettronica superficiale, tanto che gli elettroni che si trovano nel livello energetico più esterno sono spesso detti elettroni di legame.
Esistono 2 modi fondamentali attraverso cui un atomo può utilizzare tali elettroni per legarsi con un altro atomo, dando luogo ad altrettanti tipi di legami: il legame ionico ed il legame covalente.
Poichè, come abbiamo già detto, gli elettroni coinvolti nei legami chimici sono quelli che occupano il livello energetico più superficiale (elettroni di valenza), introduciamo un metodo semplice per rappresentare i primi 8 elettroni, noto come configurazione di Lewis degli elementi.
Secondo tale metodo i 2 + 6 elettroni dei primi due sottolivelli (1 orbitale s e 3 orbitali p) del livello più esterno vengono rappresentati come punti o coppie di punti disposte ai quattro lati del simbolo chimico dell'elemento. Ogni lato rappresenta un orbitale e può contenere al massimo 2 elettroni. Per maggior chiarezza diamo la configurazione di Lewis degli elementi appartenenti al 3° periodo.
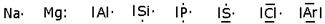
Le coppie di elettroni vengono più spesso rappresentate con una barretta.
Legame ionico
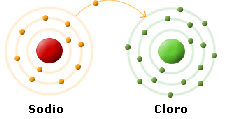
Prendiamo in considerazione un elemento appartenente al primo gruppo, ad esempio il Sodio (Na). Il Sodio ha numero atomico Z = 11 e quindi oltre a possedere 11 protoni possiede 11 elettroni. Gli 11 elettroni sono sistemati: 2 a saturare il primo livello energetico; 2 + 6 = 8 a saturare il secondo livello energetico; 1 elettrone inizia il riempimento del terzo livello energetico.
Si noti che il Sodio presenta un elettrone in più rispetto al gas nobile che lo precede nella tabella periodica, il Neon. Come sappiamo la configurazione dei gas nobili è particolarmente stabile (regola dell'ottetto) ed il Sodio tenderà dunque a liberarsi del suo elettrone superficiale per assumere la configurazione elettronica stabile del Neon.
In tal modo però viene "scoperta" una carica positiva del nucleo. Infatti il Sodio viene ad avere 11 protoni e 10 elettroni.
Il Sodio si trasforma in un atomo con una carica netta positiva o ione positivo Na+
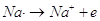
Si dicono ioni gli atomi elettricamente carichi, positivamente o negativamente. Uno ione positivo o catione si forma per perdita di uno o più elettroni.
Allo stesso modo il Calcio (Ca), che appartiene al secondo gruppo, presenta 2 elettroni in più del gas nobile che lo precede, l'Argon. Il Calcio ha un'elevata tendenza a perdere questi due elettroni per raggiungere la configurazione elettronica stabile dell'Argon.
In questo modo si trasforma in un catione con due cariche positive Ca++ o, il che è lo stesso, Ca2+ (calcio due più).
Prendiamo in considerazione ora un elemento del settimo gruppo come il Cloro con Z = 17.
Il Cloro sistema i suoi 17 elettroni: 2 nel 1° livello (saturandolo) , 8 nel 2° livello (saturandolo) ed i rimanenti 7 nel 3° livello.
Al Cloro manca un solo elettrone per raggiungere la configurazione stabile (8 elettroni) del gas nobile che lo segue (Ar) nella tabella periodica, l'Argon. Per il Cloro è infatti più semplice (energeticamente più vantaggioso) acquistare un elettrone, piuttosto che perderne 7 per raggiungere la configurazione del gas nobile che lo precede (Ne).
Acquistando 1 elettrone il Cloro si trasforma in uno ione negativo Cl-. Con un elettrone in più infatti il Cloro viene a possedere 18 elettroni contro i 17 protoni del suo nucleo, evidenziando in tal modo una carica netta negativa.
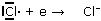
Gli ioni negativi o anioni sono atomi che hanno acquistato uno o più elettroni.
Se dunque noi avviciniamo del Cloro gassoso Cl2 e del Sodio metallico Na, assisteremo ad una violenta reazione esoergonica. Il Sodio, che presenta una elevata tendenza a liberarsi del suo elettrone, lo cede al Cloro che presenta da parte sua una grande tendenza ad acquistarlo.
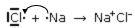
Ma lo ione Sodio Na+ e lo ione Cloro Cl- non rimangono a questo punto separati. Presentando due cariche di segno opposto essi si attirano formando una molecola, complessivamente neutra, la cui formula bruta è NaCl, il Cloruro di Sodio.
Si è dunque formato un legame chimico di tipo elettrostatico tra ioni aventi carica opposta: il legame ionico. I composti che presentano questo tipo di legame si dicono composti ionici.
Dire che entrambi gli atomi raggiungono in tal modo una configurazione più stabile significa affermare che diminuiscono il loro contenuto energetico. L'energia si libera in genere sotto forma di calore di reazione.
2Na + Cl2 ð 2 NaCl + 196,53 kcal
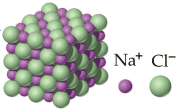
In realtà non esiste una singola molecola di Cloruro di Sodio, come abitualmente e convenzionalmente si scrive. Infatti l'attrazione tra cariche di segno opposto, come sono cationi e anioni, non si sviluppa solo in un'unica direzione, ma agisce uniformemente in tutte le direzioni (con simmetria sferica). Uno ione Na+ risulta infatti circondato da 6 ioni Cl- e viceversa., formando uno sconfinato reticolato cubico, in cui ioni di carica opposta si alternano ordinatamente nelle tre direzioni dello spazio. Tale disposizione ordinata è detta cristallina, poichè genera macroscopicamente un cristallo che conserva la geometria della sottostante struttura atomica.
Nei composti ionici quindi la formula bruta non descrive una struttura molecolare autonoma, ma indica il rapporto numerico esistente nel cristallo tra ioni positivi e negativi. Nel caso del Cloruro di Sodio, ad esempio, la formula bruta NaCl ci informa che il rapporto numerico tra ioni Na+ e Cl- all'interno del reticolo è di 1:1.
Se la reazione fosse avvenuta tra il Calcio ed il Cloro, il Calcio avrebbe ceduto due elettroni a 2 atomi di Cloro, ciascuno dei quali avrebbe acquistato un elettrone.
In tal caso affinchè il reticolato ionico sia nel complesso neutro è necessario che per ciascun ione Ca2+ siano presenti 2 ioni Cl-.
La formula CaCl2 indica dunque che nel reticolo cristallino del Cloruro di Calcio il rapporto tra ioni Calcio e ioni Cloro è 2:1.
Le sostanze ioniche, poste in acqua si dissociano negli ioni costituenti.
In acqua quindi le sostanze ioniche non sono presenti come molecole, ma come ioni.
In tutte le soluzioni acquose che costituiscono gli esseri viventi e nelle cellule in particolare, si trovano ioni di interesse biologico quali appunto: K+, Na+, Ca2+, Mg2+, Cl- etc.
Legame covalente
In generale gli ultimi elementi di ciascun periodo hanno tutti una grande tendenza ad acquistare elettroni, mentre i primi elementi di ciascun periodo presentano una forte tendenza a perdere elettroni. Quando atomi di questo tipo interagiscono tendono a scambiarsi elettroni per formare legami ionici.
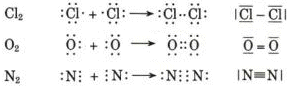
Ma come possiamo spiegare il legame esistente tra due atomi che, trovandosi alla fine di un periodo hanno entrambi una forte tendenza ad acquistare elettroni? Come spieghiamo ad esempio il legame che tiene uniti due atomi di Cloro nella molecola biatomica Cl2?
Quando due atomi di Cloro si avvicinano ciascuno dei due tende a strappare l'elettrone all'altro, senza peraltro riuscirvi.

Ma l'attrazione reciproca di ciascun nucleo sull'elettrone dell'altro atomo agisce come una forza di legame che mantiene i due atomi uniti. I due atomi di Cloro “condividono” una coppia di elettroni e tale “condivisione” costituisce il legame covalente. La condivisione avviene tramite sovrapposizione e “fusione” dei due orbitali atomici che contengono ciascuno un singolo elettrone con formazione di un unico orbitale molecolare di legame.
In questo modo ora i due elettroni non appartengono più all'uno o all'altro atomo, ma ruotano entrambi intorno all'intera struttura molecolare biatomica.
Si dice che i due elettroni sono stati messi in comune o in compartecipazione.
Ciascun nucleo "vede" ora intorno a sè i 6 elettroni non condivisi più i 2 elettroni condivisi per un totale di 8 elettroni. La condivisione di una coppia di elettroni permette a ciascun atomo di Cloro di raggiungere la configurazione stabile dell’ottetto. Alla molecola biatomica del Cloro viene assegnata la seguente formula di struttura
Cl - Cl
dove il trattino indica i due elettroni condivisi e quindi il legame covalente tra i due atomi di idrogeno.
Prendiamo ora in considerazione la molecola biatomica dell'Ossigeno O2. L'Ossigeno presenta 8 elettroni di cui 6 nel livello energetico più esterno. Quando due atomi di ossigeno si avvicinano ciascuno cerca di strappare all'altro due elettroni, senza riuscirvi, per raggiungere la configurazione dell'ottetto.
Il risultato è che in questo caso vengono messe in comune 2 coppie di elettroni con sovrapposizione di 2 coppie di orbitali e formazione di 2 orbitali di legame, ciascuno contenente una coppia di elettroni. Ciascun atomo di ossigeno ora "vede" intorno a sè 8 elettroni nel suo livello energetico più superficiale. La molecola di O2 è quindi tenuta insieme da un legame covalente doppio. La formula di struttura è
O = O
dove il doppio trattino indica i due legami.
Esistono anche legami covalenti tripli come nel caso dell'Azoto gassoso N2. Qui ciascun atomo di Azoto, possedendo solo 5 elettroni superficiali è costretto a condividerne 3 con un altro atomo di azoto per completare l'ottetto.
N ºN
I legami covalenti sono direzionali, nel senso che essi formano tra loro angoli ben determinati. Ad esempio l'angolo compreso tra i due legami Idrogeno-Ossigeno nell'acqua è di circa 105°.
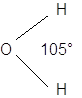
Inoltre mentre i legami semplici permettono la libera rotazione degli atomi intorno all'asse di legame, i legami doppi e tripli non permettono rotazioni. La possibilità o meno di effettuare delle torsioni interne alle molecole ha delle conseguenze notevoli soprattutto per le grosse molecole organiche. Le proteine, ad esempio, macromolecole formate da migliaia di atomi, sono in grado di assumere strutture e funzioni specifiche proprio tramite rotazioni interne attorno agli assi di legame.
Legame covalente polare
Quando gli elettroni vengono condivisi da atomi del medesimo elemento, ciascun atomo li attrae con la medesima intensità. In questo caso gli elettroni condivisi (elettroni di legame) possono essere immaginati come una nuvola negativa che si dipone in maniera omogenea e simmetrica intorno ai due nuclei senza produrre alcun tipo di polarità sulla molecola. Si parla in questo caso di legame covalente puro.
Nella maggior parte dei casi però gli atomi che formano il legame covalente appartengono ad elementi diversi che presentano una diversa forza di attrazione sugli elettroni di legame.
Si definisce elettronegatività χ (chi) la forza con cui un atomo attira a sé gli elettroni condivisi. I valori di elettronegatività si trovano tabulati nella tabella periodica e presentano il valore minimo in basso a sinistra (Francio = 0.7) e crescono diagonalmente fino ad assumere il valore massimo in alto a destra (Fluoro = 4) In genere si usa la scala di elettronegatività di Pauling.
Dunque, quando si forma un legame covalente tra due atomi che presentano diversa elettronegatività la nube elettronica che costituisce il legame covalente risulta spostata verso l'atomo più elettronegativo. Quest'ultimo acquista pertanto una carica parzialmente negativa (indicata con d-), mentre l'altro una carica parzialmente positiva (d+).La distribuzione asimmetrica della nuvola elettronica produce dunque due poli aventi carica opposta (dipòlo) ed il legame viene perciò definito covalente polare.
E’ ciò che accade, ad esempio nella molecola dell’acido Cloridrico (HCl) in cui l’Idrogeno condivide un elettrone con il Cloro. Il Cloro più elettronegativo presenta una parziale carica negativa e la molecola di HCl risulta polare
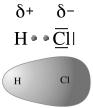
Maggiore è la differenza di elettronegatività (Δχ) tra i due elementi e maggiore sarà la polarità del legame (le cariche parziali saranno più vicine ad una intera carica).
Quando la differenza di elettronegatività tra i due elementi supera il valore critico di 1.9, si assume che l’elemento più elettronegativo sia in grado di strappare l’elettrone all’altro elemento ed il legame viene descritto come ionico.
Possiamo dunque descrivere il legame ionico come un caso limite del legame covalente polare per Δχ > 1.9
Prendiamo ora in considerazione la molecola dell'acqua H2O
Ciascun atomo di Idrogeno condivide il suo elettrone con uno dei due elettroni condivisi dall'Ossigeno
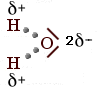
Si formano due legami covalenti. Ma le nuvole elettroniche ad essi associate vengono in questo caso attirate con maggior forza dal nucleo dell'ossigeno (più elettronegativo), il quale contiene ben 8 protoni contro il solo protone che costituisce ciascun nucleo di Idrogeno.
Ancora una volta possiamo rappresentarci la situazione immaginando che la nuvola elettronica negativa sia deformata, più densa sopra l'elemento più elettronegativo.
La carica che si forma alle estremità o poli della molecola sarebbe intera e non parziale solo se la differenza di elettronegatività tra i due elementi fosse talmente elevata da permettere all'elemento più elettronegativo di strappare l'elettrone di legame. In tal caso si formerebbero però due ioni ed il legame non verrebbe descritto più come covalente, ma come ionico.
Tutte le sostanze chimiche che al loro interno presentano dei dipoli si dicono polari, le altre apolari.
La presenza di un dipolo sulle molecole d’acqua può essere facilmente rilevata osservando che un sottile filo d'acqua che scende da un rubinetto viene deviato dalla verticale se gli viene avvicinato un bastoncino di plastica o vetro caricato elettricamente per strofinio.
Legami intermolecolari
Le molecole polari sono soggette ad ulteriori legami deboli intermolecolari, di reciproca attrazione elettrostatica, tra i poli opposti delle molecole stesse, detti legami dipolo-dipolo (o interazioni dipolo-dipolo).
Legame Idrogeno
I legami Idrogeno (o ponti a Idrogeno) sono un caso particolare di interazione dipolo-dipolo. Si presentano quando l’Idrogeno si lega con un legame covalente polare ad uno dei tre elementi più elettronegativi della tabella periodica: il Fluoro, l’Ossigeno o l’Azoto (sigla FON). In questo caso la differenza di elettronegatività e di conseguenza la polarità del legame sono così elevate da generare delle interazioni dipolo-dipolo particolarmente intense. I legami idrogeno si rappresentano come linee tratteggiate che collegano il polo positivo di un dipolo al polo negativo di un altro. In un legame Idrogeno gli atomi di idrogeno fanno da ponte tra due atomi del gruppo FON, avendo con uno di questi un legame covalente polare e con l’altro un legame elettrostatico.
In una soluzione di acqua H2O ed ammoniaca NH3, ad esempio, si formano ponti ad idrogeno tra Ossigeno ed Azoto, tra Ossigeno ed Ossigeno e tra Azoto ed Azoto.

I legami Idrogeno sono interazioni fondamentali per le strutture biologiche.
In particolare essi influenzano l'attività dell'acqua come liquido organico e sono responsabili della stabilità e del comportamento biologico di molte grosse molecole organiche (proteine e acidi nucleici).
Di tale secondo aspetto ci occuperemo quando affronteremo lo studio dei principali composti di interesse biologico, mentre ora approfondiremo il comportamento e le caratteristiche chimiche dell'acqua.
Peculiarità chimico-fisiche e importanza biologica dell'acqua
L'acqua costituisce un dipolo particolarmente intenso in quanto la differenza di elettronegatività tra Ossigeno ed Idrogeno risulta piuttosto elevata. In queste condizioni le molecole d'acqua allo stato liquido tendono a disporsi in modo da presentare il polo negativo (Ossigeno) di una rivolto verso il polo positivo (idrogeno) di un'altra. Si formano così legami intermolecolari di natura elettrostatica (legami idrogeno) che mantengono fra loro unite le molecole.
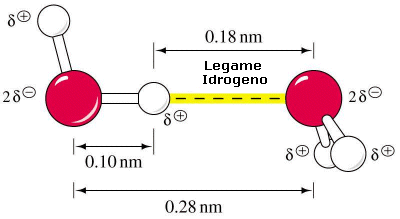
Tale legame è meno intenso di un normale legame covalente e a temperatura ambiente non è in grado di bloccare le molecole in una posizione determinata. Se ciò avvenisse l'acqua solidificherebbe.
Ciononostante il legame tra le molecole dipolari dell'acqua è molto più intenso della maggior parte dei legami esistenti tra altre molecole polari. Nell'acqua i legami a idrogeno si formano e si spezzano in continuazione (si calcola che la vita media di un legame a idrogeno sia di circa 10-11 s) lasciando libere le molecole di muoversi scivolando le une rispetto all'altro, ma garantendo complessivamente una notevole forza di coesione tra esse.
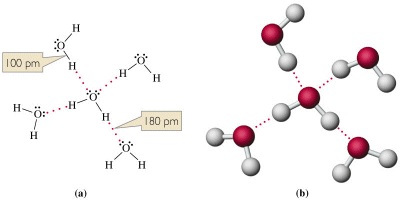
La presenza del legame a idrogeno tra le molecole d'acqua è fondamentale per comprendere le caratteristiche chimico-fisiche e biologiche di quest'ultima.
La presenza dell'acqua è essenziale per qualsiasi forma di vita conosciuta. E' nell'acqua che si ritiene sia nata la vita e di acqua sono costituite, per gran parte della loro massa, le cellule.
Temperatura e passaggi di stato |
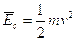 , dove k è la costante di Boltzmann. Ciò significa che a temperature più elevate corrispondono energie cinetiche maggiori e quindi maggiori velocità delle molecole. Possiamo rappresentarci una sostanza allo stato solido come costituita di particelle (atomi, ioni o molecole) che vibrano intorno a posizioni di equilibrio ben determinate senza poter sostanzialmente mutare la loro posizione reciproca. Aumentando la temperatura tali particelle vibrano sempre più rapidamente finchè si separano e, passando allo stato liquido, sono in grado di muoversi le une rispetto alle altre, pur rimanendo aderenti.
, dove k è la costante di Boltzmann. Ciò significa che a temperature più elevate corrispondono energie cinetiche maggiori e quindi maggiori velocità delle molecole. Possiamo rappresentarci una sostanza allo stato solido come costituita di particelle (atomi, ioni o molecole) che vibrano intorno a posizioni di equilibrio ben determinate senza poter sostanzialmente mutare la loro posizione reciproca. Aumentando la temperatura tali particelle vibrano sempre più rapidamente finchè si separano e, passando allo stato liquido, sono in grado di muoversi le une rispetto alle altre, pur rimanendo aderenti.
Caratteristiche termiche dell'acqua e funzione termoregolatrice
L'acqua presenta un elevato calore specifico.
Il calore specifico di una sostanza è il numero di calorie necessarie per elevare di 1°C la temperatura di un grammo di sostanza. Ricordando la definizione di caloria è evidente che il calore specifico dell’acqua è 1 cal/(g °C).
Dire che l’acqua presenta un elevato calore specifico significa che, a parità di massa, l'acqua necessita di una quantità di calore molto più elevata di qualsiasi altra sostanza inorganica (ammoniaca (NH3) esclusa) per manifestare lo stesso effetto di riscaldamento (lo stesso aumento di temperatura). In altre parole l'acqua, rispetto ad altre sostanze può assorbire molto calore senza scaldarsi eccessivamente. Una conseguenza di questo fenomeno è che l'acqua si scalda (e si raffredda) molto più lentamente delle altre sostanze.
L'elevata capacità termica dell'acqua (quantità di calore necessaria per aumentare di 1°C la temperatura di un corpo di massa qualsiasi) si spiega con la presenza dei legami idrogeno tra le sue molecole. Quando forniamo calore all'acqua solo una parte di questo si trasforma in un aumento di energia cinetica delle molecole, e quindi in un aumento di temperatura, poichè in parte esso è utilizzato per spezzare i legami idrogeno che imbrigliano le strutture molecolari.
Per lo stesso motivo l'acqua assorbe anche molto calore durante il processo di evaporazione, presenta cioè un elevato calore di evaporazione (a 25°C e 1 Atm 0,584 kcal/g). Ancora una volta il calore fornito deve vincere delle forze di attrazione tra le molecole particolarmente intense (legami idrogeno) per farle passare allo stato di vapore, abbandonando la superficie del liquido.
L'elevata capacità termica e l'elevato calore di evaporazione fanno dell'acqua un ottimo liquido refrigerante e, in generale, termoregolante, capace cioè di ammortizzare in modo molto efficace gli sbalzi termici.
E' ad esempio un fatto noto che l'acqua del mare si scalda e si raffredda più lentamente della terraferma e quindi le grandi masse d'acqua presenti sulla superficie terrestre, come un'enorme volano termico, riducono le escursioni termiche giornaliere e stagionali, svolgendo un'azione mitigatrice sul clima delle località marine e lacustri.
Ma al di là delle modificazioni climatiche, gli effetti più importanti da questo punto di vista, l'acqua li manifesta sugli esseri viventi. L'acqua presente negli organismi (dal 50% al 95% in peso) è infatti in grado di assorbire in modo particolarmente efficace le variazioni esterne di temperatura. In particolare alcuni esseri viventi, tra cui l'uomo, sono in grado di eliminare calore in eccesso facendo evaporare acqua (sudorazione).
Il mantenimento dell'equilibrio termico è un fatto vitale per gli esseri viventi in quanto molte macromolecole sono termolabili e vengono distrutte ad elevate temperature, mentre a basse temperature la velocità delle reazioni chimiche diminuisce al punto tale da essere incompatibile con la vita.
Un'altra caratteristica notevole dell'acqua è che quando solidifica aumenta di volume, contrariamente a quanto fanno la maggior parte delle altre sostanze chimiche. Ciò è dovuto al fatto che la solidificazione avviene attraverso una stabilizzazione dei legami idrogeno, i quali costringono ora le molecole a disporsi in una struttura cristallina ordinata, a maglie più larghe rispetto a quanto avveniva nel liquido. Se il ghiaccio fosse più pesante dell'acqua si accumulerebbe, anno dopo anno, sul fondo dei bacini idrici senza possibilità di ritornare allo stato liquido. In tal modo le riserve idriche mondiali si esaurirebbero (diventando tra l'altro sempre più salate) ed ogni forma vivente in esse sarebbe destinata all'estinzione. La formazione di strati di ghiaccio galleggiante tende invece a proteggere l'acqua sottostante da ulteriori diminuzioni di temperatura.
Capillarità e imbibizione
Per la sua elevata polarità l'acqua aderisce (forze di adesione) fortemente a qualunque superficie costituita da molecole che presentino cariche elettriche. In tal modo si spiega perchè l'acqua "bagna". Tale capacità dell'acqua è alla base dei fenomeni di capillarità ed imbibizione, per i quali l'acqua è in grado di salire attraverso tubi sottili (capillari) o di invadere materiali porosi, letteralmente scalando le loro superfici. I fenomeni di capillarità ed imbibizione presentano una notevole valenza biologica, soprattutto nel mondo vegetale, in quanto sono alla base della circolazione dei fluidi nelle piante. L'imbibizione è ad esempio fondamentale per i processi di germinazione, poichè permette di spaccare i resistenti tegumenti che proteggono il seme (si ritiene che gli Egiziani staccassero i blocchi di pietra necessari per costruire le piramidi, praticando dei fori all'interno dei quali facevano dilatare dei bastoni di legno per imbibizione).
Le soluzioni acquose ed il pH
Ma l'azione biologica probabilmente più importante svolta dall'acqua è quella di agire da mezzo disperdente per le molecole.
Tutte le reazioni chimiche che avvengono all'interno della cellula richiedono infatti che i reagenti siano intimamente mescolati a livello molecolare. Il sistema che ne deriva prende il nome di soluzione acquosa. In una soluzione acquosa l'acqua è il solvente, mentre le sostanze presenti in concentrazione minore sono i soluti. Le reazioni che avvengono in soluzione sono molto più veloci di reazioni di tipo diverso, poiché essendo i reagenti completamente mescolati a livello molecolare hanno una maggior probabilità di scontrarsi e reagire.
Inoltre le sostanze disciolte in acqua possono essere trasportate all'interno dell'organismo in modo semplice ed efficiente (sangue, linfa etc).
L'acqua non scioglie qualsiasi sostanza, ma solo le sostanze dotate di carica elettrica (ioniche e polari) per le quali presenta grande affinità. Infatti i dipoli che costituiscono le molecole d'acqua vengono attirati dagli ioni o dalle molecole polari. L'attrazione che l'acqua esercita su tali particelle (interazioni idrofile) è talmente elevata da arrivare a vincere le forze interne di coesione. Le molecole di soluto vengono quindi staccate ad una ad una, rivestite di molecole d'acqua (idratazione) e portate in soluzione.
Nel caso ad esempio di un soluto ionico, come il Cloruro di Sodio, gli ioni Na+ vengono circondati da molecole d'acqua che puntano verso di essi il proprio polo negativo (Ossigeno), mentre gli ioni Cl- attirano i poli positivi delle molecole d'acqua (Idrogeno).
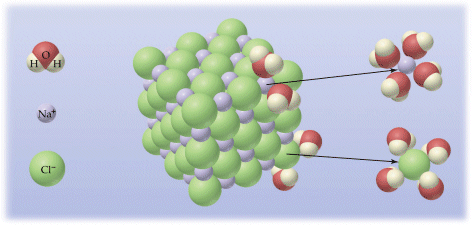
Se il soluto è costituito di particelle di dimensioni superiori a 10-3 m (1 m = 10-3 mm) e inferiori a 1 m, (sopra il micron si parla di sospensioni) la soluzione presenta delle caratteristiche particolari e viene detta soluzione (o sistema) colloidale. Nei colloidi il soluto prende il nome di fase dispersa. A differenza di quanto avviene nelle soluzioni vere, nei colloidi è possibile evidenziare le particelle della fase dispersa tramite l'effetto Tyndall, illuminando cioè il colloide ed osservandolo perpendicolarmente alla direzione del fascio luminoso. Le particelle della fase dispersa, avendo dimensioni dell'ordine di grandezza della lunghezza d'onda della luce impiegata per illuminarle, la diffondono e diventano così visibili (è lo stesso effetto che si produce quando un raggio di sole che entra da una finestra si “materializza” illuminando il pulviscolo atmosferico).
Inoltre se aumentiamo la concentrazione di una soluzione vera il soluto si deposita come corpo di fondo cristallino, mentre in un sistema colloidale ciò non avviene ed esso o diventa gelatinoso o coagula (floccula) formando grossi agglomerati irregolari (albume d'uovo, sangue etc). I colloidi diluiti sono simili alle soluzioni vere e sono detti sol, quelli più concentrati presentano un aspetto gelatinoso e sono detti gel.
Le soluzioni biologiche, oltre a presentare soluti di piccole dimensioni (ioni, zuccheri, amminoacidi etc), contengono anche grosse macromolecole (soprattutto proteine) che ne fanno dei sistemi che presentano sia le caratteristiche delle soluzioni vere che quelle delle soluzioni colloidali.
L'acqua non scioglie le sostanze apolari come i grassi (lipidi). Le molecole apolari vengono letteralmente escluse dall'acqua, per cui esse tendono a formare grossi aggregati tra di loro (olio in acqua). Tali effetti di esclusione a carico di molecole apolari vanno sotto il nome di interazioni idrofobiche.
Nelle cellule le interazioni idrofobiche hanno grande importanza, sia nel costituire riserve di sostanze lipidiche, sia soprattutto nel determinare la struttura delle membrane cellulari. Le membrane che circondano tutte le cellule e le separano dall'ambiente esterno sono in pratica costituite da sottili strati lipidici stabilizzati dalle repulsioni idrofobiche che l'acqua esercita sia dall'interno che dall'esterno della cellula.
Il comportamento dell'acqua è legato infine ad un'altra importante caratteristica delle soluzioni: l'acidità.
Nell'acqua pura una piccolissima percentuale di molecole (circa 2 molecole ogni miliardo) si dissociano secondo la reazione
H2O Û H+ + OH-
In pratica un atomo di Idrogeno si separa dalla molecola d'acqua alla quale apparteneva, abbandonando il suo elettrone. L'Idrogeno si distacca quindi come ione positivo o idrogenione, mentre lo ione negativo che si forma prende il nome di ione ossidrile o idrossido (OH-).
Naturalmente nell'acqua pura il numero di ioni H+ è uguale al numero di ioni OH-. In tali condizioni la soluzione si dice neutra.
Ma se introduciamo nell'acqua un soluto in grado di rilasciare ioni H+ o ioni OH- l'equilibrio viene alterato.
Le sostanze che in acqua sono in grado di dissociarsi, facendo aumentare gli ioni H+ sono dette acide. Un esempio è l'acido cloridrico (HCl), una sostanza secreta anche dal nostro stomaco
HCl Û H+ + Cl-
Quando in una soluzione vi è un eccesso di ioni H+ rispetto agli ioni OH-, la soluzione si dice acida
Le sostanze che in acqua sono in grado di dissociarsi, facendo aumentare gli ioni OH- sono dette basiche. Un esempio è l'idrossido di sodio (NaOH)
NaOH Û Na+ + OH-
Quando in una soluzione vi è un eccesso di ioni OH- rispetto agli ioni H+, la soluzione si dice basica Per misurare la concentrazione degli ioni H+ si ricorre ad una scala convenzionale, la scala del pH
pH = 7 soluzione neutra
pH < 7 soluzione acida
pH > 7 soluzione basica
Una soluzione è tanto più acida quanto più basso è il suo pH ed è tanto più basica quanto più elevato il suo pH. La scala del pH va solitamente da 0 a 14, ma sono possibili anche valori esterni a tale intervallo. La scala del pH è una scala logaritmica. Ciò significa che per ogni variazione di un punto del pH la concentrazione degli ioni H+ varia di un fattore 10. Una soluzione a pH = 2 è 104 = 10.000 più concentrata in ioni H+ di una soluzione a pH = 6
In genere ogni soluzione biologica presenta un pH ben determinato e non tollera variazioni eccessive.
Il sangue umano ad esempio presenta un pH = 7,4 ed una variazione superiore a ± 0,3 può rapidamente portare al coma ed alla morte. Questa enorme sensibilità alle variazioni di pH è essenzialmente legata al fatto che le proteine che controllano il nostro metabolismo (enzimi) lavorano correttamente solo a certi valori di pH. Ad esempio la pepsina, un enzima che inizia la digestione delle proteine nello stomaco, è in grado di lavorare solo con pH compresi tra 1 e 2 (questo è uno dei motivi per cui le pareti dello stomaco sono in grado di secernere grandi quantità di acido cloridrico).
La chimica della vita
Da un punto di vista chimico ciò che distingue un essere vivente dalla materia inanimata non è il tipo di elementi chimici di cui è costituito, ma la complessità con cui tali elementi si legano per dare origine alle enorme varietà delle sostanze organiche.
Il Carbonio che costituisce il diamante, la grafite e le rocce calcaree è assolutamente identico al carbonio che forma l'emoglobina del sangue o il glucosio, ma in tali molecole biologiche esso si lega a formare strutture incomparabilmente più complesse.
Gli esseri viventi richiedono in definitiva composti chimici altamente organizzati, ordinati e spesso caratterizzati da un elevato contenuto energetico.
Le molecole biologiche sono composte peraltro da un numero relativamente esiguo di elementi chimici. Dei 92 elementi esistenti in natura gli esseri viventi ne utilizzano in pratica una trentina e quasi tutti a basso peso atomico (i primi elementi della tabella periodica).
Tra questi alcuni sono presenti in tracce, altri vengono utilizzati in maniera massiccia, basti pensare che il 99% di un essere vivente è formato da 6 elementi chimici: il Carbonio (C), l'Idrogeno (H), l'Ossigeno (O), l'Azoto (N), il Fosforo (P) e lo Zolfo (S). [sigla CHONPS].
Il carbonio
Un posto centrale nella chimica della vita spetta al Carbonio. Il motivo principale è che il Carbonio, oltre ad essere un elemento particolarmente diffuso in natura, possiede 4 elettroni superficiali che può condividere con altri elementi chimici (ed anche con altri atomi di Carbonio) per formare quattro legami covalenti stabili.
Il fatto più interessante è però la capacità del carbonio di legarsi con altri atomi di carbonio per formare lunghe catene, variamente ramificate o addirittura chiuse a formare anelli (in realtà poligoni), mantenendo nel contempo legami liberi che vengono saturati da altri elementi chimici (essenzialmente H, O, N). In tal modo le combinazioni possibili risultano virtualmente infinite.
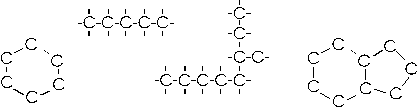
In molti casi le macromolecole organiche raggiungono complessità e varietà ancor maggiori attraverso il processo di polimerizzazione.
Un polimero è un composto formato dalla successione di molte molecole uguali (monomeri) o di pochi tipi diversi. Uno stesso monomero può agganciarsi in modi diversi formando polimeri aventi differenti caratteristiche (il glucosio, ad esempio, forma sia la cellulosa che l'amido).
Il Carbonio va quindi a costituire una specie di impalcatura, rappresenta lo scheletro di tutte le sostanze organiche, al punto che la chimica organica è conosciuta anche come chimica del carbonio.
L'unica sostanza con caratteristiche chimiche molto simili è il Silicio, l'elemento con il quale si costruiscono i chips dei computer. Forse in qualche altro mondo potrebbe esistere una vita basata sulla chimica del Silicio.
Quando il Carbonio si lega con se stesso produce molecole dalla geometria variabile che dipende dagli angoli di legame.
Si possono a tal proposito distinguere tre casi:
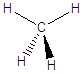
carbonio tetraedrico
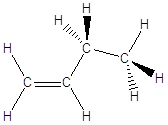
il Carbonio forma 4 legami covalenti semplici (carbonio tetraedrico)
in questo caso i quattro legami sono tra loro equidistanti e formano l'uno con l'altro angoli di legame di circa 109°. Possiamo immaginare l'atomo di Carbonio al centro di un tetraedro con i quattro legami che puntano verso i vertici.

La famiglia più semplice di composti organici che esemplifica questa geometria è quella degli alcani. Gli alcani sono degli idrocarburi saturi. Il termine “idrocarburi” sta ad indicare dei composti formati solo da idrogeno e carbonio. Il termine “saturi” sta ad indicare il fatto che la catena carboniosa è saturata dall’idrogeno, presenta cioè il numero massimo di atomi di idrogeno possibili. Gli alcani sono classificati in relazione al numero di atomi di carbonio che forma la catena carboniosa
Metano |
CH4 |
Etano |
C2H6 |
Propano |
C3H8 |
Butano |
C4H10 |
Pentano |
C5H12 |
Esano |
C6H14 |
Eptano |
C7H16 |
Ottano |
C8H18 |
Nonano |
C9H20 |
Decano |
C10H22 |
etc |
CnH2n+2 |
La desinenza che caratterizza i loro nomi è -ano
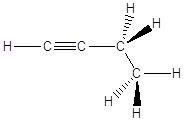
In queste condizioni le catene che si formano non hanno andamento rettilineo, ma a zig-zag. Se immaginiamo da esempio i legami Carbonio-Carbonio sul piano del foglio, i legami residui escono davanti e dietro il foglio. Per rappresentare la reciproca disposizione degli atomi e dei legami nello spazio si rappresentano i legami giacenti sul foglio come segmenti continui, i legami in avvicinamento (rispetto all’osservatore) come cunei pieni ed i legami in allontanamento come cunei tratteggiati.

Metano |
Etano |
Propano |
|
|
|

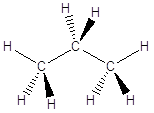
Si utilizzano anche altre rappresentazioni più compatte dei composti organici come le formule razionali e le formule schematiche
Formula razionale: la struttura viene descritta in modo sintetico condensando gli atomi di idrogeno intorno al carbonio al quale sono legati
Metano |
Etano |
Propano |
Butano |
|
CH3 – CH3 |
CH3 – CH2 – CH3 |
CH3 – CH2 – CH2 – CH3 |
Formula schematica rappresenta solo la catena carboniosa della molecola attraverso una linea spezzata. Le estremità aperte della spezzata rappresentano gruppi CH3, mentre i vertici gruppi CH2. Tutti gli altri elementi chimici (che non siano C ed H) vengono rappresentati.
Metano |
Etano |
Propano |
Butano |
|
|
|
|

Poiché i legami sono semplici gli atomi possono ruotare attorno all'asse di legame Carbonio-Carbonio e, in opportune condizioni, chiudersi ad anello.
Naturalmente non sono possibili tutte le forme geometriche. Gli angoli interni non possono infatti essere molto diversi dai 109° caratteristici del carbonio tetraedrico.
Le forme più stabili (che non producono tensioni eccessive sugli angoli di legame) e quindi di gran lunga più frequenti sono l'anello esagonale e pentagonale.
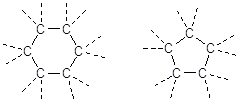
Carbonio planare
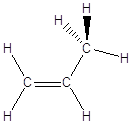
Il Carbonio forma 2 legami semplici ed 1 doppio (carbonio planare)
In questo caso i legami giacciono su di un piano a 120° l'uno dall'altro.
Non vi è libera rotazione intorno al legame Carbonio-Carbonio
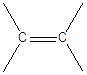
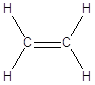
La famiglia più semplice di composti organici che esemplifica questa geometria è quella degli alcheni. Gli alcheni sono degli idrocarburi insaturi. Il termine “insaturi” sta ad indicare il fatto che la catena carboniosa non è completamente saturata dall’idrogeno. Nel caso specifico l’insaturazione si manifesta con la presenza di un doppio legame tra due atomi di carbonio. Idrogenando il doppio legame (aggiungendo due atomi di idrogeno) si può infatti trasformare un alchene in un alcano. Gli alcheni hanno la stessa radice dell’alcano con ugual numero di atomi di carbonio, ma cambiano la desinenza in –ene.
Etene |
C2H4 |
Propene |
C3H6 |
Butene |
C4H8 |
Pentene |
C5H10 |
Esene |
C6H12 |
Eptene |
C7H14 |
Ottene |
C8H16 |
Nonene |
C9H18 |
Decene |
C10H20 |
etc |
CnH2n |
Etene |
C2H4 |
Etene |
Propene |
Butene |
|
|
|
CH2 = CH2 |
CH2 = CH – CH3 |
CH2 = CH – CH2 – CH3 |
|
|
|

Nelle formule schematiche i vertici rappresentano gruppi CH2, CH o C a seconda che vi convergano rispettivamente 2, 3 o 4 legami (segmenti).

Carbonio lineare
Il Carbonio forma 1 legame semplice ed 1 triplo oppure 2 legami doppi(Carbonio lineare). In questo caso i legami giacciono lungo una retta a 180° l'uno dall'altro
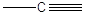
Anche in questo caso è impedita la libera rotazione intorno all'asse Carbonio-Carbonio interessato dal legame triplo.
La famiglia più semplice di composti organici che esemplifica questa geometria è quella degli alchini. Anche gli alchini, come gli alcheni, sono degli idrocarburi insaturi. Nel caso specifico l’insaturazione si manifesta con la presenza di un triplo legame tra due atomi di carbonio. Gli alchini hanno la stessa radice dell’alcano con ugual numero di atomi di carbonio, ma cambiano la desinenza in –ino.
Etino |
C2H2 |
Propino |
C3H4 |
Butino |
C4H6 |
Pentino |
C5H8 |
Esino |
C6H10 |
Eptino |
C7H12 |
Ottino |
C8H14 |
Nonino |
C9H16 |
Decino |
C10H18 |
etc |
CnH2n-2 |
Etino |
C2H2 |
Etino |
Propino |
Butino |
|
|
|
CH ≡ CH |
CH ≡ C – CH3 |
CH ≡ C – CH2 – CH3 |
|
|
|
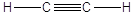
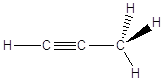

Legami semplici, doppi e tripli possono comunque succedersi in modi diversi all'interno di una stessa molecola organica. Naturalmente queste strutture carboniose possiedono sempre legami liberi che utilizzano per legarsi con altri elementi chimici. In questo modo essi forniscono alla molecola che si forma le sue caratteristiche chimiche peculiari e aumentano grandemente il numero di combinazioni possibili tra atomi.
I composti organici che costituiscono tutti gli esseri viventi possono essere raggruppati in quattro classi fondamentali: glucidi, lipidi, protidi e acidi nucleici.
Glucidi (carboidrati o zuccheri)
In natura i glucidi si trovano come zuccheri semplici o monosaccaridi e zuccheri composti da 2 o più monosaccaridi. In quest'ultimo caso si parla rispettivamente di disaccaridi e polisaccaridi.
I glucidi sono costituiti solo da tre elementi chimici: Carbonio, Idrogeno e Ossigeno.
Monosaccaridi
I monosaccaridi sono formati da catene di atomi di carbonio, costituite da un minimo di 3 ad un massimo di 7 atomi. I monosaccaridi più diffusi in natura sono quelli con sei (esosi) e con cinque (pentosi) atomi di carbonio. I pentosi e gli esosi si chiudono in genere ad anello.
I chimici classificano i composti organici in relazione alla presenza nella loro molecola di particolari gruppi chimici, detti gruppi funzionali. Ciascuna classe di composti organici presenta uno o più gruppi funzionali caratteristici.
I monosaccaridi sono caratterizzati dalla presenza di due gruppi funzionali: l'ossidrile ed il carbonile.
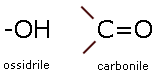
Un monosaccaride presenta il primo (aldosi) o il secondo (chetosi) carbonio della catena unito ad un atomo di ossigeno con un doppio legame covalente (gruppo funzionale carbonilico), mentre tutti gli altri atomi di carbonio sono uniti con un gruppo ossidrilico. Tutti i rimanenti legami del carbonio vengono saturati dall'idrogeno.
La desinenza -osio caratterizza i nomi dei monosaccaridi (glucosio, fruttosio etc).
Costruiamo un esoso generico con il carbonile in C1 (aldoesoso) ed in C2 (chetoesoso)
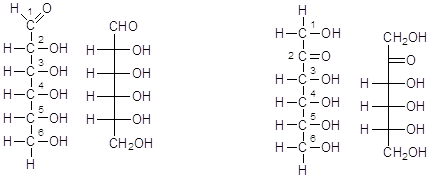
Aldoesoso Chetoesoso
Queste sono le formula di struttura di un esoso generico. Esistono infatti diversi esosi con la stessa formula bruta C6H12O6, che differiscono tra loro per la posizione dei 5 gruppi ossidrili ( a destra o a sinistra della catena carboniosa). L'esoso di gran lunga più diffuso in natura è il glucosio che presenta l'ossidrile C3 a sinistra
In realtà gli esosi si chiudono ad anello poichè il gruppo carbonilico reagisce con l’ossidrile in C5 formando un esagono (con l'ossigeno che fa da ponte tra C1 e C5).
Il glucosio, ad esempio, si chiude ad esagono
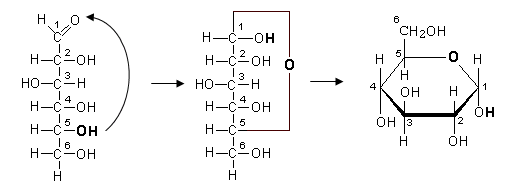
Mentre il fruttosio (chetoesoso) si chiude a pentagono

Tra gli aldoesosi più diffusi ricordiamo il galattosio (ossidrili C3 e C4 a sinistra) che si trova abbondante nel latte.
I pentosi possiedono tutti formula bruta C5H10O5.
Tra i pentosi ricordiamo il ribosio, un aldopentoso con struttura ciclica a pentagono (l'ossigeno fa da ponte tra C1 e C4). Il ribosio è una molecola fondamentale per la costruzione di molte sostanze organiche complesse, tra cui gli acidi nucleici (DNA ed RNA).
Infine tra i triosi ricordiamo l'aldeide glicerica o gliceraldeide, il più semplice tra gli zuccheri naturali, un aldotrioso che rappresenta un importante intermedio nel metabolismo degli zuccheri.
Tutti i monosaccaridi sono caratterizzati dalla formula bruta generica CnH2nOn. In altre parole C, H ed O sono sempre presenti nel rapporto di 1:2:1. Per ogni atomo di carbonio ve ne sono 2 di idrogeno ed 1 di ossigeno. Poichè l'idrogeno e l'ossigeno sono presenti nelle stesse proporzioni in cui si trovano nella molecola d'acqua (2:1), i glucidi vengono anche detti carboidrati (idrati di carbonio). Ciò non significa naturalmente che nella loro molecola esista acqua in quanto tale.
La presenza di numerosi ossidrili, caratterizzati da una elevata polarità, rende i monosaccaridi facilmente solubili in acqua
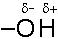
I monosaccaridi rappresentano il combustibile per eccellenza delle cellule. In particolare il glucosio che può essere velocemente trasportato e "bruciato" per fornire energia indispensabile alla vita. L'energia si trova concentrata soprattutto nei legami C-H, più energetici dei legami C-O.
Molti monosaccaridi esplicano comunque anche altre funzioni biologiche, fungendo soprattutto da mattoni chimici per la costruzione di molecole complesse.
Disaccaridi e Polisaccaridi
I monosaccaridi possono facilmente unirsi tra loro facendo reagire 2 gruppi ossidrili con perdita di una molecola d’acqua, tramite una reazione detta di condensazione, rimanendo uniti tramite un atomo di ossigeno.
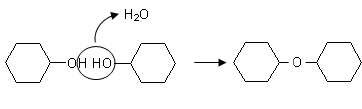
Quando si uniscono in questo modo due molecole di glucosio si forma un disaccaride detto maltosio.
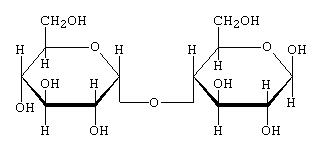
maltosio
Tra i disaccaridi ricordiamo inoltre il saccarosio (glucosio + fruttosio) e il lattosio (glucosio + galattosio).
Naturalmente la condensazione può avvenire tra molte molecole di monosaccaride formando lunghe catene polimeriche, variamente ramificate. In questo modo gli esseri viventi sintetizzano i polisaccaridi.
Certi polisaccaridi vengono sintetizzati per immagazzinare riserve di zuccheri, altri hanno invece funzioni strutturali (servono alla cellula come materiale da costruzione).
Hanno funzione di riserva di energia l'amido (nelle piante) e il glicogeno (negli animali). Entrambi sono polimeri del glucosio, ma differiscono nel modo in cui gli ossidrili reagiscono durante la condensazione. L'amido si trova abbondante nei semi e nei tuberi delle piante, mentre il glicogeno si concentra soprattutto nei muscoli e nel fegato. I polisaccaridi di riserva non possono essere utilizzati così come sono per fornire energia, ma devono essere preventivamente riconvertiti nei monosaccaridi costituenti. La reazione avviene con l'introduzione di una molecola d'acqua nel punto in cui era stata precedentemente eliminata. La molecola d'acqua spezza il ponte ossigeno riformando i due ossidrili. Tale processo di separazione attraverso l'acqua è detto idrolisi.
Nelle piante alcuni polisaccaridi presentano importanti funzioni strutturali. Il più importante tra essi è certamente la cellulosa, un polimero del glucosio utilizzato dalle cellule vegetali per formare la parete cellulare, un rivestimento protettivo esterno alla membrana cellulare. La maggior parte degli animali non possiede l'enzima necessario per idrolizzare il legame che tiene unite le molecole di glucosio nella cellulosa. I mammiferi erbivori sono in grado di utilizzare in parte il glucosio della cellulosa grazie alla presenza nel loro apparato digerente di batteri simbionti capaci di idrolizzarla.
Negli esseri viventi animali è più raro trovare polisaccaridi con funzioni strutturali. Possiamo ricordare comunque la chitina, un polisaccaride costituito da un derivato azotato del glucosio (acetilglucosammina) utilizzato dai crostacei e dagli insetti per costruire i loro esoscheletri.
Lipidi
I lipidi sono una classe abbastanza eterogenea di composti chimici aventi come caratteristica principale la quasi totale insolubilità in acqua. Gran parte dei legami chimici presenti nelle molecole lipidiche sono infatti legami C-H, i quali sono praticamente apolari e quindi idrofobici.
Un lipide immerso in ambiente acquoso forma una fase separata proprio in virtù delle interazioni idrofobiche.
Per queste loro caratteristiche i lipidi vengono utilizzati dagli esseri viventi come materiale isolante ed impermeabilizzante.
Inoltre, poiché i legami C-H sono particolarmente energetici, i lipidi costituiscono generalmente un'ottima riserva di energia, facilmente accumulabile in depositi per la loro insolubilità. A parità di peso un lipide è in grado di fornire più del doppio di energia rispetto ad un carboidrato (circa 9 kcal/g contro le 3,75 kcal/g di un monosaccaride e le 4 kcal/g di un polisaccaride o di una proteina).
Il contenuto di energia di un composto chimico per unità di peso può essere descritto tramite il suo grado di ossidazione o di riduzione.
Una reazione di ossidazione è una reazione in cui un composto chimico perde elettroni ed energia, mentre una riduzione è una reazione in cui un composto chimico acquista elettroni ed energia. Poiché nei composti organici nella maggior parte dei casi una ossidazione avviene per sostituzione di un legame C-O con un legame C-H, possiamo giudicare il grado di ossidazione o di riduzione di un composto organico dal numero di tali legami. In prima approssimazione possiamo dunque affermare che un composto sarà tanto più ossidato quanto più numerosi sono i suoi legami C-O e sarà tanto più ridotto quanto più numerosi sono i suoi legami C-H.
Così ad esempio un atomo di carbonio legato a 4 atomi di idrogeno a formare una molecola di metano (CH4) è un composto molto energetico, con un elevato grado di riduzione (non per nulla lo usiamo come combustibile).
Quando bruciamo il metano per estrarne l'energia noi effettuiamo un'ossidazione. Sostituiamo infatti gli atomi di idrogeno con gli atomi di ossigeno ottenendo un composto estremamente ossidato e povero di energia, l'anidride carbonica CO2. La differenza di energia tra i due composti è quella che noi sfruttiamo.
Ma tra il metano che presenta un elevato grado di riduzione e l'anidride carbonica che risulta completamente ossidata esistono composti chimici che presentano gradi di ossidazione intermedi.
Ad esempio se sostituiamo un idrogeno del metano con un ossidrile, otteniamo un composto parzialmente ossidato, ma ancora ricco di energia, l'alcool metilico o metanolo (CH3OH).
Gli alcoli sono una famiglia di composti caratterizzati dal gruppo funzionale ossidrile (-OH) e dalla desinenza -olo (ricordiamo che i composti organici si riuniscono in famiglie caratterizzate dalla presenza di un particolare raggruppamento chimico detto gruppo funzionale)
Se ossidiamo ulteriormente il metanolo otteniamo l'aldeide formica o formaldeide o metanale (H-CHO). Le aldeidi sono caratterizzate dal gruppo funzionale aldeidico (-CHO) e presentano desinenza -ale.
Continuando nel processo di ossidazione otteniamo un composto ancora più povero di energia, l'acido formico o acido metanoico (H-COOH). Gli acidi organici (o carbossilici) sono caratterizzati dal gruppo funzionale carbossile (-COOH) e dalla desinenza -oico.
Ossidando l'acido formico si ottiene infine l'anidride carbonica.
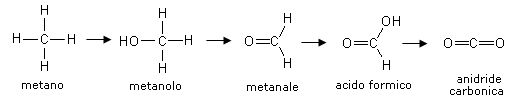
Come si può notare man mano che procede il processo ossidativo gli atomi di idrogeno vengono sostituiti da atomi di ossigeno. Inoltre per molti composti il grado di ossidazione è relativo. Infatti diremo che l'aldeide formica è più ossidata dell'alcool metilico, ma è più ridotta dell'acido formico.
Ritorniamo ora ai lipidi. Da un punto di vista chimico un lipide è formato dall'unione di un acido carbossilico con un alcool ed è quindi un estere.
Indichiamo un acido generico come R-COOH ed un alcool generico come R-OH. La lettera R indica un radicale generico, un gruppo di atomi in catena carboniosa di cui, nel caso particolare, non interessa la composizione.
L'acido e l'alcol si uniscono utilizzando i rispettivi gruppi funzionali, tramite una reazione di condensazione detta esterificazione. Durante la reazione viene eliminata una molecola d'acqua tra l'ossidrile del gruppo carbossilico e l'idrogeno del gruppo alcolico.
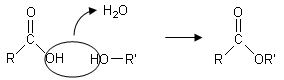
Anche i lipidi possono essere idrolizzati per ottenere gli acidi e gli alcoli di partenza.
I lipidi sono un sottoinsieme degli esteri, nel senso che gli esseri viventi utilizzano solo alcuni tipi di alcoli ed alcuni tipi di acidi organici (acidi grassi) per sintetizzare i lipidi.
Negli acidi grassi il gruppo R è costituito da lunghe catene di atomi di carbonio (fino a 22) saturate da atomi di idrogeno
CH3 CH2 CH2 COOH acido butirrico
CH3 (CH2)16 COOH acido stearico
Alcuni acidi grassi contengono doppi legami tra atomi di carbonio lungo la catena. Ciascun doppio legame è dovuto alla mancanza di due atomi di idrogeno. Per questo motivo tali acidi grassi sono detti insaturi e, se vi è più di un doppio legame, polinsaturi. Un'esempio di acido insaturo è l'acido oleico
CH3 (CH2)7 CH = CH (CH2)7 COOH acido oleico
Un acido polinsaturo essenziale per l'alimentazione umana (il nostro organismo è incapace di sintetizzarlo) è l'acido linolenico
CH3 (CH2 CH = CH )3 (CH2)7 COOH acido linolenico
I lipidi si dividono in lipidi semplici (formati solo dall'unione di alcool e acido grasso) e lipidi composti (formati anche da una terza sostanza chimica).
Lipidi semplici
I lipidi semplici si dividono in trigliceridi, ceridi (o cere) e steridi (o lipidi steroidei). Ciascuno di questi tre gruppi è caratterizzato da un particolare alcol.
I trigliceridi sono caratterizzati dalla presenza dell'alcol glicerolo (o glicerina), un alcool triossidrilico

Glicerolo
Per inciso se ossidiamo il glicerolo nel carbonio C1, otteniamo l'aldeide glicerica.
Facendo reagire i tre gruppi alcolici con tre acidi grassi, con perdita di tre molecole d'acqua si ottiene un trigliceride
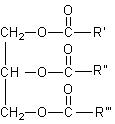
I 3 acidi grassi possono essere dello stesso tipo o o di tipo diverso.
I trigliceridi possono essere saturi o insaturi.
Sono saturi quando gli acidi grassi costituenti hanno tutti i legami saturati da atomi di idrogeno. Sono insaturi quando una o più coppie di atomi di carbonio adiacenti formano doppi legami a causa dell'assenza dell'idrogeno.
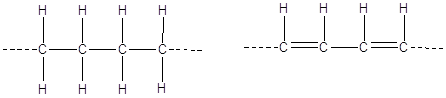
saturo insaturo
Ciò che distingue i vari acidi grassi tra loro, oltre alla lunghezza della catena, è il numero e la posizione dei doppi legami.
I trigliceridi sono composti fondamentali nella dieta umana e si ritrovano come componenti essenziali negli esseri viventi, dove costituiscono i principali lipidi di riserva e di deposito.
Quelli di derivazione vegetale sono in genere insaturi o polinsaturi. In queste condizioni essi sono liquidi a temperatura ambiente e vengono definiti olii.
Quelli di derivazione animale (lardo, burro etc) sono saturi e per questo motivo sono solidi a temperatura ambiente. Vengono definiti grassi.
Attraverso processi chimici è possibile saturare i trigliceridi insaturi (idrogenazione) ottenendo dei grassi vegetali spalmabili (margarina).
I ceridi, o semplicemente cere. sono caratterizzati dalla presenza di un alcool monoossidrilico a lunga catena carboniosa.
Ad esempio il costituente principale della cera d'api è il palmitato di miricile, formato dalla reazione dell'acido palmitico (C15H31COOH) con l'alcool miricilico (C31H63OH).
La cera carnauba, che si ricava dalle foglie di una palma sudamericana, è formata essenzialmente di cerotato di miricile (acido cerotico C25H51COOH + alcool miricilico).
Le cere sono in genere utilizzate da piante ed animali per rendere impermeabili i loro rivestimenti esterni (le penne degli uccelli, la superficie esterna di molte foglie e frutti), per proteggere e mantenere morbidi e flessibili peli e cute.
Gli steridi o lipidi steroidei utilizzano come alcol uno sterolo. Tra gli steroli, il più importante è il colesterolo.
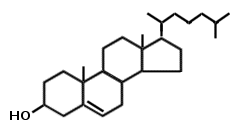
COLESTEROLO
Il colesterolo si trova nel nostro organismo sia libero (25 - 40 %) che esterificato da acidi grassi a lunga catena. In forma esterificata è uno dei costituenti principali degli steridi.
Il colesterolo è una molecola indispensabile al nostro organismo essendo un componente della membrana ed il precursore di importanti ormoni. Tuttavia si ritiene che un livello eccessivo di colesterolo nel sangue (ipercolesterolemia) costituisca un rischio di aterosclerosi, depositandosi in placche (ateromi, atheros = pericoloso, athera = poltiglia?) sulle pareti dei vasi sanguigni, diminuendone il lume e aumentando in tal modo il rischio di malattie cardiovascolari (infarti, ictus).
Il colesterolo viene in parte introdotto nell'organismo attraverso la dieta ed in parte sintetizzato dal fegato a partire dagli acidi grassi saturi. Il fegato è anche l'organo che demolisce il colesterolo in eccesso. Nel sangue il colesterolo viene trasportato da due tipi di proteine andando a formare i complessi LDL (Low Density Lipoprotein = a basso peso molecolare) ed HDL (High Density Lipoprotein = ad alto peso molecolare).
Le LDL trasportano il colesterolo alle cellule per il suo utilizzo. Le cellule presentano sulla loro membrana esterna dei recettori in grado di riconoscere la frazione proteica delle LDL. Dopo aver agganciato le LDL la membrana cellulare si introflette risucchiandole all'interno (endocitosi mediata da recettori). Se i recettori cellulari per le LDL mancano (per un difetto genetico) o sono alterati il colesterolo ematico è destinato ad aumentare in modo pericoloso.
Le HDL trasportano il colesterolo in eccesso al fegato per essere demolito ed escreto (attraverso la bile) fungendo così da "spazzini". Sembra che un elevato rapporto HDL/LDL diminuisca drasticamente il rischio di malattie cardiovascolari. Per questo motivo le HDL sono comunemente note come "colesterolo buono" e le LDL come "colesterolo cattivo". Il rapporto HDL/colesterolo totale è definito indice di rischio cardiovascolare. In un soggetto sano l’indice di rischio dovrebbe essere inferiore a 5 per i maschi e a 4,5 per le femmine.
L'attività sportiva ed una dieta non troppo ricca di grassi saturi sembrano aumentare i livelli di HDL, mentre fumo e alcool li diminuiscono.
Lipidi composti
Sono formati da lipidi semplici in associazione con altre molecole.
Sono ad esempio lipidi composti le lipoproteine, proteine che trasportano i lipidi nel sangue come le HDL e le LDL o i glicolipidi, formati dall'unione di un lipide semplice (in cui l'alcol è in genere la sfingosina, un alcol amminato) con monosaccaridi, componenti essenziali delle membrane cellulari (soprattutto delle cellule nervose come i cerebrosidi, i gangliosidi ecc.).
Noi descriveremo in particolare i fosfolipidi o fosfatidi, lipidi semplici legati ad una molecola di acido fosforico (H3PO4). Nei glicerofosfatidi, ad esempio, l'acido fosforico sostituisce una delle molecole di acido grasso di un trigliceride.
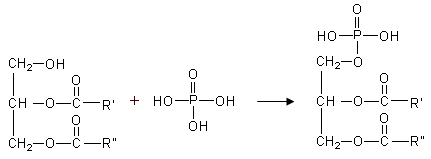
I fosfolipidi sono i costituenti fondamentali delle membrane biologiche. Ne esistono diversi tipi. Fra i più importanti sono da ricordare le lecitine e le cefaline, formati da glicerolo esterificato nelle prime due funzioni alcoliche da acidi grassi, e nella terza da una fosfatilammina, cioè un gruppo fosforico legato ad una ammina, come la colina (HOCH2CH2-N+(CH3)3) o l’etanolammina (HOCH2CH2-NH2).
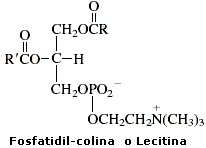
I fosfolipidi possiedono particolari caratteristiche di polarità che ne fanno dei costituenti fondamentali delle membrane cellulari (lecitine). L'acido fosforico è infatti polare mentre le due lunghe catene degli acidi grassi sono apolari. L'intera molecola viene solitamente rappresentata attraverso una "testa idrofila" che contiene il gruppo fosfato polare e due "code idrofobe", costituite dai residui degli acidi grassi apolari.

L'eteropolarità permette ai fosfolipidi di organizzarsi spontaneamente in acqua, formando strutture ordinate. Le teste tendono infatti a disporsi a contatto con l'acqua, mentre le code, respinte dalle interazione idrofobiche, tendono a restare rivolte le une verso le altre. Si possono in tal modo formare strutture micellari sferoidali e lamine bistratificate.
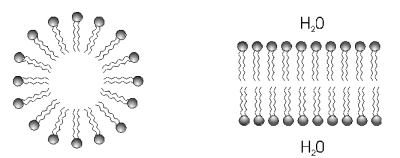
Protidi
I protidi o proteine o polipeptidi sono le sostanze chimiche che svolgono i compiti più svariati all'interno della cellula. Possiamo comunque ricondurre le funzioni proteiche all'interno di due principali attività: a) strutturali o plastiche b) di regolazione o controllo.
Vi è una stretta correlazione fra forma e funzione delle proteine.
Le proteine con funzione plastica hanno una forma fibrosa. Sono costituite da catene polipeptidiche allungate, disposte in fasci lungo uno stesso asse a costituire le fibre. Sono insolubili in acqua.
Le proteine con funzione di regolazione e controllo hanno invece una forma globulare. Le catene sono strettamente avvolte in forma compatta, sferica o globulare, come un gomitolo. Sono solubili in acqua.
Esiste un numero enorme di proteine, una diversa dall'altra, sia all'interno di uno stesso organismo, sia tra organismi di specie diverse, ma tutte sono ottenute attraverso la combinazione di 20 mattoni chimici: gli amminoacidi. Il fatto che le proteine siano diverse e caratteristiche per ogni individuo e per ogni specie è legato al fatto che esse vengono sintetizzate a partire dalle informazioni genetiche contenute nel DNA
Oltre ai venti amminoacidi comuni a tutte le proteine di tutti gli esseri viventi, ve ne sono alcuni specifici di alcune proteine (l'ossiprolina del collagene) e alcuni che svolgono da soli azioni fisiologiche particolari (ormone tiroxina).
I 20 amminoacidi comuni a tutti gli esseri viventi si legano a formare lunghe catene proteiche, ognuna costituita da qualche centinaio di amminoacidi.
Il nostro organismo non è in grado di sintetizzare 8 amminoacidi, per questo detti amminoacidi essenziali, che devono essere pertanto introdotti con la dieta.
Tali amminoacidi sono particolarmente abbondanti nelle proteine di origine animale che, per questo motivo, sono dette "nobili", rispetto a quelle di origine vegetale.
Tutti gli amminoacidi che costituiscono le proteine presentano la seguente struttura generale
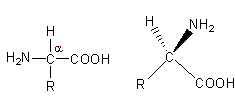
Un carbonio centrale tetraedrico (detto carbonio alfa) al quale è legato un gruppo amminico (-NH2) di natura basica, un gruppo carbossilico (-COOH) di natura acida, un atomo di idrogeno ed un gruppo chimico (-R), detto residuo amminoacidico o gruppo R, diverso da amminoacido ad amminoacido.
Gli amminoacidi si legano tra loro a formare le proteine tramite un legame di condensazione, facendo reagire il gruppo amminico di un amminoacido con il gruppo carbossilico di un altro, con perdita di una molecola di acqua.
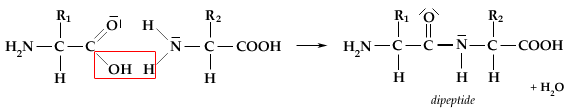
Il legame che si produce è detto legame peptidico ed il gruppo chimico CONH è detto gruppo peptidico.
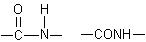
gruppo peptidico
In questo modo tutte le proteine si presentano costituite da un lungo filamento chimico comune, formato dalla successione di gruppi CH e CONH, dal quale sporgono i residui amminoacidici (-R), la cui successione è diversa da proteina a proteina. Tale successione definisce la peculiare struttura primaria di una proteine.
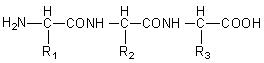
Convenzionalmente la struttura primaria si rappresenta iniziando con il gruppo amminico e terminando con il gruppo carbossilico. Il primo amminoacido della sequenza è detto amminoacido N-terminale (enne-terminale o ammino-terminale), l'ultimo amminoacido C-terminale (ci-terminale o carbossi-terminale).
Sequenze di pochi amminoacidi (50 - 100) sono dette peptidi. Oltre un certo limite (diverso da autore ad autore: PM » 5.000 - 10.000 uma) si parla di polipeptidi.
I filamenti proteici non rimangono mai lineari. Rispondendo alle sollecitazioni prodotte dalle loro polarità interne si ripiegano su se stessi formando strutture a diverso grado di complessità. Sono stati descritti per le proteine 4 livelli strutturali principali.
- La struttura primaria è costituita dalla successione degli amminoacidi
- La struttura secondaria è definita dai tipi di avvolgimenti (stabilizzati da ponti idrogeno) che interessano tratti del filamento proteico (Eliche, Foglietti e Curve)
- La struttura terziaria è definita dal modo in cui il filamento proteico si ripiega su se stesso in una conformazione compatta di tipo globulare contenente le diverse strutture secondarie
- La struttura quaternaria si genera quando diverse molecole proteiche in struttura terziaria globulare si saldano tra loro, assemblandosi
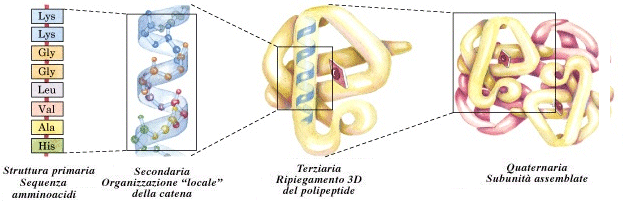
Fonte: http://rexmagister.altervista.org/downloads/BiologiaDOC.zip
sito web: http://rodomontano.altervista.org/biologia.php
Autore del testo: Rodomontano
Biologia appunti parte 1
Visita la nostra pagina principale
Biologia appunti parte 1
Termini d' uso e privacy