Biologia appunti parte 4
Biologia appunti parte 4
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Eredità poligenica: caratteri non mendeliani a variabilità continua
La maggior parte dei caratteri degli individui sono caratteri non mendeliani e manifestano pertanto una variabilità continua e non discreta. Si tratta in genere di caratteri tipicamente quantitativi come il peso, l’altezza, la forma del naso etc. Anche quando si tratta di caratteri numerabili (numero dei capelli) il numero delle classi numeriche è così elevato che la variabilità può comunque essere considerata continua.
L’ereditarietà di questi caratteri è più complessa rispetto ai caratteri mendeliani che si presentano in poche forme alternative ed in proporzioni caratteristiche. In particolare questi caratteri non si prestano ad essere selezionati. Se infatti si tenta di selezionare un particolare valore di un carattere continuo (ad esempio una pianta alta 1,50 cm all’interno di piante che presentano tutte le altezze comprese tra uno e due metri) segliendo nella progenie solo gli individui con quel valore ed incrociandoli, si ottengono sempre anche tutte le altre gradazioni.
Questo comportamento (non selezionabilità) aveva indotto i genetisti dell’inizio del ‘900 a ritenere che la variabilità di questi caratteri non avesse una base genetica, ma solo ambientale. Gli esperimenti condotti nei primi decenni del ‘900 dimostrarono che, oltre alla componente ambientale, tali caratteri presentano una base genetica di tipo poligenico, sono cioè caratteri controllati da numerosi geni (eredità poligenica).
Tutti i geni che controllano un carattere a variabilità continua presentano solo due tipi di alleli, detti fattori. Per questo motivo l’eredità poligenica è nota anche come eredità polifattoriale. I fattori possono essere solo di due tipi: fattori positivi (+) che spingono la manifestazione del carattere verso un estremo del suo intervallo di variabilità e fattori negativi (-) che spingono la manifestazione del carattere verso l’altro estremo del suo intervallo di variabilità. La manifestazione del carattere dipenderà dunque dalla particolare combinazione di fattori positivi e negativi.
I caratteri a variabilità continua presentano inoltre una particolare distribuzione di frequenza, nota come curva a campana o gaussiana.
Se ad esempio poniamo in ascissa le altezze della popolazione umana ed in ordinata la frequenza con cui gli individui di una determinata altezza compaiono nella popolazione, troveremo

Gli individui che presentano i valori estremi del carattere (con tutti i fattori positivi o tutti i fattori negativi) sono molto rari, mentre avvicinandoci ai valori intermedi gli individui diventano sempre più frequenti.
Incrociando individui che si trovano agli estremi opposti dell’intervallo di variabilità si ottiene una progenie di individui con caratteri intermedi (regressione filiale). Incrociando gli individui intermedi la generazione successiva tende nuovamente a distribuirsi secondo una gaussiana.
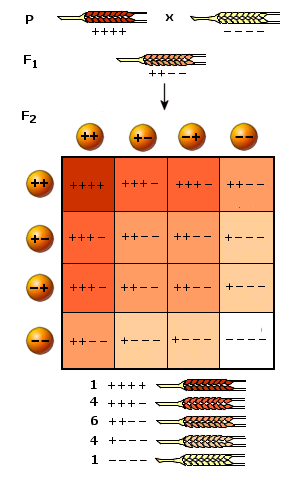
Un esempio particolarmente semplice di eredità poligenica è rappresentato dal colore della cariosside del frumento, controllato da due soli geni. Il colore della cariosside dipende dalla quantità di pigmento presente e va dal rosso cupo al bianco attraverso 5 tonalità complessive di colore.
Se incrociamo due individui che stanno alle due estremità opposte dell’intervallo di variabilità, una pianta a semi rosso cupo (tutti e 4 i fattori positivi: genotipo ++ ++) con una pianta a semi bianchi (tutti e 4 i fattori negativi -- --) otterremo una pianta che riceverà ovviamente un gamete ++ da un genitore ed un gamete –- dall’altro. In F1 avremo pertanto un individuo dal colore intermedio, con due fattori positivi e due negativi (++ --).
Incrociando la progenie F1 e ipotizzando che i due geni possano ricombinarsi liberamente (assortimento indipendente) essi produrranno 4 tipi di gameti (++, +-, -+, --) che, posti ai due lati di un quadrato di Punnett, daranno 16 individui.
1 con quattro fattori positivi ++++
4 con tre fattori positivi ed uno negativo +++-
6 con due fattori positivi e due negativi ++--
4 con un fattori positivo e tre negativi +---
1 con quattro fattori negativi ----
Se riportiamo le 5 classi fenotipiche in un istogramma che ne evidenzi la distribuzione di frequenza, avremo

Come si può osservare due geni sono troppo pochi per ottenere una curva continua, ma l’aspetto è gia quello della distribuzione gaussiana.
Un carattere controllato da tre geni (6 fattori) presenta in F2 7 classi di frequenza. Per costruire la distribuzione di frequenza dovremmo incrociare un individuo con tutti e 6 i fattori positivi (++ ++ ++) con un individuo con tutti e 6 i fattori negativi (-- -- --). Ovviamente otterremo in F1 individui intermedi con tre fattori positivi e tre negativi (+- +- +-). Se incrociamo tra loro gli individui intermedi, nell’ipotesi che i fattori siano indipendenti e che quindi durante la gametogenesi vi sia una completa ricombinazione tra i fattori positivi e negativi, avremo 23 = 8 tipi di gameti (+++, ++-, +-+, +--, ---, --+, -+-, -++). Posizionando i gameti ai due lati di un quadrato di Punnett otterremo 8 x 8 = 64 combinazioni genotipiche.
È comunque possibile evitare di riempire il quadrato di Punnett e trovare la distribuzione di frequenza cercata, sviluppando la potenza pari di un binomio
(P + N)f
Dove
P = fattore positivo
N = fattore negativo
f = numero di fattori
Per 3 geni e quindi 6 fattori si ottiene
(P + N)6 = 1P6N0 + 6P5N1 + 15P4N2 + 20P3N3 + 15P2N4 + 6P1N5 + 1P0N6
i 7 monomi che si ottengono corrispondono alle 7 classi di frequenza, con i coefficienti dei monomi che corrispondono agli individui di ciascuna classe.
1P6N0 |
1 individuo con 6 fattori positivi |
1 |
++ ++ ++ |
6P5N1 |
6 individui con 5 fattori positivi ed 1 negativo |
6 |
++ ++ +- |
15P4N2 |
15 individui con 4 fattori positivi e 2 negativi |
15 |
++ ++ -- |
20P3N3 |
20 individui con 3 fattori positivi e 3 negativi |
20 |
++ +- -- |
15P2N4 |
15 individui con 2 fattori positivi e 4 negativi |
15 |
++ -- -- |
6P1N5 |
6 individui con 1 fattore positivo e 5 negativi |
6 |
+- -- -- |
1P0N6 |
1 individuo con 6 fattori negativi |
1 |
-- -- -- |

è evidente che al crescere del numero di fattori che controllano il carattere il numero di classi di variabilità aumenterà e l’istogramma sarà sempre più simile ad una curva.
Mutazioni genetiche
Le mutazioni sono alterazioni casuali, permanenti ed ereditarie dell’informazione genetica di una cellula a livello della struttura o dell’organizzazione del suo DNA.
Il DNA è suscettibile di danneggiamento
- ad opera di agenti chimici e fisici presenti nell’ambiente (agenti mutageni)
- a causa del normale metabolismo cellulare che produce intermedi reattivi (radicali liberi) in grado di attuare modificazioni ossidative sulle molecole organiche
- a causa di errori nei processi cellulari di divisione cellulare durante la duplicazione del DNA o durante i processi meiotici di crossing-over e separazione degli omologhi.
Essendo modificazioni casuali a carico di un’informazione altamente strutturata ed ordinata, le mutazioni hanno una bassissima probabilità di produrre una alterazione favorevole dell’organismo. È un po’ come se dessimo in mano i circuiti di un televisore ad una scimmia. La probabilità che alla fine riesca a trasformarli in quelli di un computer è praticamente nulla.
La maggior parte delle mutazioni sono dunque eventi sfavorevoli per l’individuo in cui si manifestano. Alcune di esse possono essere anche cancerogene (o teratogene), trasformando la cellula che le subisce in una cellula tumorale. In alcuni casi possono essere addirittura letali. Si deve comunque tener presente che le conseguenze per un organismo di una mutazione dannosa sono diverse a seconda che si manifestino a carico delle cellule somatiche o delle cellule germinali (riproduttive).
Se una cellula mutata dà origine solo a cellule somatiche (non sessuali) verrà prodotta un’area o un settore mutato ma queste nuove caratteristiche non verranno trasmesse alle generazioni successive. Questo tipo di mutazione è chiamata mutazione somatica.
Se, al contrario, la mutazione avviene nella linea germinale (cellule sessuali o riproduttive) di organismi che si riproducono sessualmente, la mutazione potrà essere trasmessa alla generazione successiva dando origine ad un individuo mutato sia nelle sue cellule somatiche sia nella linea germinale. Queste mutazioni vengono chiamate mutazioni della linea germinale.
Tuttavia, in una prospettiva biologica più ampia rispetto a quella del singolo individuo, le mutazioni sono considerate un evento fondamentale e positivo per l’evoluzione biologica. Le mutazioni sono infatti le principali responsabili della comparsa di nuovi geni e di nuovi alleli. In questo modo le mutazioni creano nuova variabilità genetica sulla quale può agire la selezione naturale. In assenza di mutazioni una specie non potrebbe evolvere ed avrebbe una elevata probabilità di non adattarsi alle modificazioni ambientali e conseguentemente di estinguersi.
Il valore biologico delle mutazioni non è dunque univoco: è in genere negativo per il singolo individuo, mentre è positivo per la specie (dal che si deduce che gli interessi del singolo individuo non coincidono necessariamente con gli interessi della specie alla quale appartiene).
Gli agenti mutageni fisici sono il calore, le radiazioni elettromagnetiche ad alta energia (raggi UV, raggi X e raggi gamma) e le radiazioni corpuscolari (radiazione alfa (nuclei di elio), radiazione beta (elettroni), neutroni e protoni). I mutageni fisici degradano il DNA, spezzandone i legami chimici.
Gli agenti mutageni chimici si classificano in analoghi dei nucleotidi, modificatori ed intercalanti.
Gli analoghi dei nucleotidi (o analoghi delle basi) sono molecole che presentano una somiglianza chimica con le normali base azotate, ma che la cellula non riconosce come diverse ed incorpora nel nuovo DNA che sintetizza.
Gli agenti modificatori (alchilanti, idrossilanti, deaminanti) alterano la struttura molecolare delle basi causando appaiamenti errati
Gli agenti intercalanti (acridine, bromuro di etidio) si inseriscono fra le basi azotate, legandosi ad esse e causano inserzioni e delezioni durante la replicazione del DNA.
Le mutazioni genetiche si classificano in geniche, cromosomiche e genomiche.
Mutazioni geniche
Le mutazioni geniche alterano un singolo gene, modificando una singola base azotata. Poiché avvengono in un punto preciso del gene sono anche dette mutazioni puntiformi.
Si classificano in mutazioni geniche per sostituzione, inserzione e delezione.
Le mutazioni geniche per sostituzione producono la sostituzione di una base azotata (ed ovviamente anche della sua complementare), modificando una tripletta di basi che codifica per un amminoacido. Gli effetti possono essere diversi. Infatti la nuova tripletta potrà codificare per il medesimo amminoacido, per nessun amminoacido o per un amminoacido diverso.
Nel caso la nuova tripletta codifichi per il medesimo amminoacido (sappiamo che più triplette possono codificare per il medesimo amminoacido), la sostituzione non avrà alcun effetto sulla struttura primaria della proteina (sequenza degli amminoacidi) e la mutazione si definisce mutazione silente.
Nel caso la nuova tripletta non codifichi per nessun amminoacido (tripletta STOP), la proteina mutata si spezza in due frammenti e perde ovviamente la sua funzionalità. La mutazione è definita mutazione non-senso.
Nel caso la nuova tripletta codifichi per un amminoacido diverso la sostituzione viene definita mutazione di senso (missense) ed i suoi effetti sulla funzionalità proteica dipendono dal punto in cui è avvenuta la mutazione. In una proteina possiamo infatti individuare dei tratti funzionali, la cui alterazione comprometterebbe l’attività della molecola, e dei tratti non funzionali in cui la sostituzione di un amminoacido non produce effetti.
Le sostituzioni amminoacidiche in tratti funzionali possono dunque alterare in modo più o meno grave la funzionalità proteica. Ne è un esempio l’anemia falciforme, una malattia genetica recessiva, in cui i globuli rossi assumono una caratteristica forma a falce. L’anemia falciforme è dovuta alla sostituzione di un unico amminoacido (valina al posto dell'acido glutammico) nelle catene beta dell’emoglobina.
Le sostituzioni amminoacidiche in tratti non funzionali sono dette mutazioni neutre. Tali mutazioni non portano infatti ne’ vantaggi ne’ svantaggi, poiché lasciano inalterata la funzionalità proteica. Per questo motivo le mutazioni neutre non sono “viste” dalla selezione naturale e possono pertanto accumularsi con ritmo costante ed in punti casuali all’interno delle regioni non funzionali di una proteina.
Le mutazioni neutre sono alla base degli orologi molecolari che i biologi utilizzano per ricostruire gli alberi filogenetici dell’evoluzione.
Un orologio molecolare è una proteina che si trova in molte specie diverse (proteina ubiquitaria), ad esempio l’emoglobina nei mammiferi o il citocromo c utilizzato nella catena respiratoria. Confrontando dunque la struttura primaria di una medesima proteina che si trova in due specie differenti (ad esempio l’emoglobina del cavallo con quella del cane) si possono evidenziare il numero di mutazioni neutre accumulatesi in termini di amminoacidi sostituiti. Una volta tarato l’orologio molecolare (cioè una volta stimato il tempo medio in milioni di anni affinché si accumuli una mutazione neutra), è possibile trasformare il numero di mutazioni trovate nel tempo trascorso dal momento in cui viveva l’antenato comune delle due specie studiate.

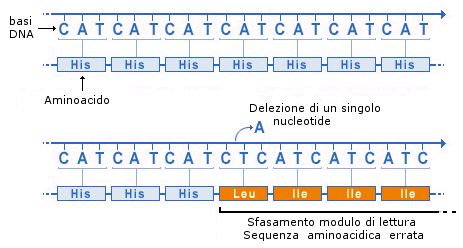
Le mutazioni geniche per inserzione provocano l’inserimento di un nuovo nucleotide.
Le mutazioni geniche per delezione provocano l’eliminazione di un nucleotide.
Queste mutazioni provocano oltre ad una modificazione del significato della tripletta di nucleotidi mutata, anche una sfasatura nella lettura di tutte le triplette successive. Queste mutazioni sono note come mutazioni con slittamento del modulo di lettura (frame-shift). Il cambiamento di tutti gli amminoacidi successivi al punto in cui è avvenuta la mutazioni porta per lo più alla perdita della funzionalità proteica.
Mutazioni cromosomiche
Le mutazioni cromosomiche (o aberrazioni) sono alterazioni della struttura dei cromosomi che interessano uno o più geni e che si producono per lo più come conseguenza di errori durante il crossing-over, tramite rotture ed errate ricomposizioni dei cromosomi. In genere, i cromosomi rotti tendono a ricomporre le rotture e a ripristinare le strutture originarie, ma a volte, può capitare che il processo di riparazione non risulti corretto e si originano cromosomi anomali con strutture nuove. Si possono, così, verificare perdite, duplicazioni, inversioni o trasferimenti di pezzi più o meno estesi di cromosoma. I geni si vengono così a trovare in una posizione diversa da quella originale. Dato che la regolazione dell’attività di un gene dipende, in parte, anche dalla sua localizzazione nel genoma, le mutazioni cromosomiche hanno, generalmente, effetti estremamente drammatici.
Le mutazioni cromosomiche si classificano in inversioni, traslocazioni, duplicazioni e delezioni.
L’inversione è una aberrazione strutturale cromosomica (riarrangiamento di struttura) che origina da due rotture sul cromosoma, successiva rotazione di 180° del tratto compreso tra le rotture e sua reintegrazione nel cromosoma stesso. Le inversioni possono essere pericentromeriche (comprendono la regione del centromero e le rotture si verificano sul braccio corto e sul braccio lungo del cromosoma) e paracentromeriche (non comprendono la regione del centromero e le due rotture si verificano sullo stesso braccio cromosomico). L’inversione si origina in genere durante l’appaiamento meiotico degli omologhi con formazione di un’ansa di inversione (loop).

La traslocazione (trasposizione) è una aberrazione cromosomica strutturale che consiste nel trasferimento di un segmento di cromosoma in una posizione errata, con un cambiamento nella localizzazione di un segmento cromosomico.
La traslocazione può essere intracromosomica o intercromosomica.
La prima implica un cambiamento di posizione di un tratto cromosomico entro lo stesso cromosoma, sia da un braccio cromosomico all’altro sia entro lo stesso braccio.
La seconda implica lo spostamento di un segmento cromosomico ad un altro non omologo.
La traslocazione intercromosomica si definisce reciproca se ciascuno dei due cromosomi non omologhi trasferisce all’altro un proprio segmento.
La traslocazione intercromosomica si definisce non reciproca se il trasferimento è unidirezionale e solo uno dei due cromosomi trasferisce all’altro un proprio segmento.
Una traslocazione non reciproca tra cromosomi omologhi produce una delezione su di un cromosoma (quello che perde il segmento) ed una duplicazione sull’altro cromosoma (quello che riceve il segmento).
Le traslocazioni bilanciate sono il caso più fortunato: consistono infatti nello “scambio alla pari” di frammenti fra cromosomi diversi. Questo tipo di traslocazione non comporta perdita di materiale genetico e perciò i portatori di una traslocazione bilanciata non manifestano in genere alcun segno clinico. Nel caso di una traslocazione non bilanciata, uno o più cromosomi in seguito alla traslocazione hanno subito la perdita di materiale genetico, mentre altri ne hanno in sovrappiù. Chi è portatore di una traslocazione bilanciata, pur non manifestando alcun sintomo, rischia di avere figli portatori di traslocazioni patologiche (non bilanciate).
La delezione è una aberrazione cromosomica strutturale che porta alla perdita di un segmento di cromosoma. In alcuni casi è associata alla duplicazione, poiché il segmento reciso viene trasferito sul cromosoma omologo dove si trovano i medesimi alleli. In altri casi il segmento viene semplicemente perso. La delezione si produce in genere con una modalità analoga all’inversione, con formazione di un’ansa di delezione che, invece di risaldarsi in posizione invertita, si stacca.
Le delezioni si possono evidenziare in un cariotipo attraverso l’assenza di una o più bande cromosomiche, o addirittura di un intero braccio, come avviene ad esempio nella sindrome 18p (p indica il braccio corto di un cromosoma, q il braccio lungo), dove è avvenuta la completa delezione del braccio corto di uno dei due cromosomi 18. L’effetto di una delezione dipende dalla grandezza della porzione mancante e dal tipo di informazione genetica in essa contenuta: in una sola banda cromosomica possono trovarsi centinaia di geni. Le microdelezioni, rispetto alle delezioni, consistono nella perdita di frammenti cromosomici più piccoli e non sono evidenziabili attraverso un normale cariotipo.
La duplicazione è una aberrazione cromosomica strutturale che porta alla presenza di due segmenti genici identici all’interno di un medesimo cromosoma. La duplicazione è in genere sempre accompagnata dalla contemporanea delezione del segmento duplicato nel cromosoma omologo (il segmento viene infatti trasferito durante il crossing-over da un omologo all’altro).

Eventi mutazionali quali delezioni, inversioni o traslocazioni possono avvicinare o allontanare sequenze nucleotidiche precedentemente contigue e pertanto modificare anche drasticamente l'espressione di geni coinvolti in tali riarrangiamenti cromosomici. Il diverso tipo di espressione di un gene in funzione della posizione occupata dal gene stesso all'interno del cromosoma e dei rapporti con geni limitrofi è definito effetto di posizione.
Esempi di cariotipi anomali e loro interpretazione |
|
Cariotipo |
interpretazione |
47,XX,+21 |
Cariotipo femminile (XX) a 47 cromosomi (47), per la presenza di un cromosoma 21 sovrannumerario (+) (Sindrome di Down) |
47,XY,+21 |
Cariotipo maschile (XY) con cromosoma 21 sovrannumerario (+) |
45,X |
Cariotipo con monosomia del cromosoma X, solitamente associato alla sindrome di Turner |
47,XXX |
Cariotipo con trisomia del cromosoma X |
47,XXY |
Cariotipo con costituzione XXY dei cromosomi del sesso, associato alla sindrome di Klinefelter |
46,XY,t(1;3)(p32;q12) |
Cariotipo maschile in cui è presente una traslocazione (t) coinvolgente un cromosoma 1 ed un cromosoma 3. I punti di rottura della traslocazione sono localizzati a livello della banda 32 sul braccio corto (p) del cromosoma 1 ed a livello della banda 12 del braccio lungo (q) del cromosoma 3. |
46,XY, del(5p) |
Cariotipo maschile in cui è presente una delezione (del) del braccio corto (p) di un cromosoma 5. (Sindrome “cri-du-chat”) |
46,XX, del(1)(p32.2) |
Cariotipo femminile in cui è presente una delezione (del) di parte del braccio corto (p) di un cromosoma 1, a livello della regione 3, nella banda 2, sottobanda 2. |
46,XY, inv(7)(p13;q22) |
Cariotipo maschile in cui è presente una inversione (inv) del cromosoma 7 del tratto che va dalla banda 13 del braccio corto (p) alla banda 22 del braccio lungo (q) |
46,XY, dup(2)(q11;q21) |
Cariotipo maschile in cui è presente una duplicazione (dup) del tratto che va dalla banda 11 alla banda 21 del braccio lungo (q) del cromosoma 2 |
Mutazioni genomiche
Le mutazioni genomiche sono alterazioni del numero dei cromosomi, come conseguenza della perdita o dell’aggiunta di interi cromosomi (cromosomi soprannumerari). Le mutazioni genomiche sono per lo più causate da una mancata disgiunzione degli omologhi durante la meiosi. Durante la gametogenesi può infatti capitare che i cromosomi omologhi non si distribuiscano equamente nei gameti. Un gamete con una anomalia di questo tipo, con cromosomi in difetto o in eccesso rispetto al suo normale corredo aploide, fecondato da un gamete normale, darà luogo ad uno zigote che non presenterà il suo normale corredo diploide. Quando una cellula presenta il suo corredo cromosomico normale, aploide o multiplo di esso, si definisce euploide. La variazione della condizione euploide con variazione del numero corretto di cromosomi si definisce aneuploidia.
Le aneuploidie sono classificate da alcuni autori come aberrazioni cromosomiche.
La monosomia e la trisomia sono esempi di aneuploidia.
La monosomia è la presenza di un singolo cromosoma al posto di una coppia all’interno di un corredo diploide.
La trisomia è la presenza di tre omologhi al posto di una coppia all’interno di un corredo diploide.
Se la mancata disgiunzione riguarda l’intero corredo cromosomico si possono formare cellule polipoidi (triploidi 3n, tetraploidi 4n etc) con comparsa di intere serie di cromosomi soprannumerari. La poliploidia è in genere una condizione letale per gli animali, mentre nei vegetali è piuttosto diffusa (frumento, cotone, tabacco) e può produrre gigantismo e maggior produttività.

Il numero atipico di cromosomi può interessare sia i cromosomi sessuali che i restanti (autosomi) . Le anomalie degli autosomi comportano effetti fenotipici molto più gravi rispetto a quelli riconducibili ai cromosomi sessuali. La maggior parte dei feti con gravi anomalie degli autosomi non arriva alla nascita in quanto il loro sviluppo è talmente anormale che la gravidanza si risolve precocemente con aborto spontaneo. Nell’ambito delle anomalie cromosomiche, molto comune fra gli aborti naturali è la poliploidia.
Aneuploidie autosomiche
Le anomalie degli autosomi, diversamente da quelle dei cromosomi sessuali, comportano effetti talmente gravi che sono compatibili con la vita solo la trisomia 21, responsabile della sindrome di Down, la trisomia 13, responsabile della sindrome di Patau e la trisomia 18, responsabile della sindrome di Edwards. Le altre trisomie complete non sono compatibili con la vita e quando sono presenti in un feto sono causa di aborto spontaneo.
La sindrome di Down o trisomia 21 è causata dalla mancata disgiunzione dei cromosomi della coppia 21 in tre copie. Le persone con tale sindrome hanno in tutte le loro cellule 47 cromosomi con il cromosoma 21. L’incidenza della trisomia aumenta drasticamente con l’età della madre. Gli affetti presentano palpebre anomali, naso schiacciato, lingua larga, mani corte e robuste, statura bassa, ritardo mentale, probabili malformazioni cardiache, attesa di vita minore del normale.
La sindrome di Patau o trisomia 13 è caratterizzata da gravi ritardi mentali. L’incidenza è più bassa rispetto alla sindrome di Down, ma come questa è legata all’età della madre. Gli affetti manifestano malformazioni fisiche come cranio ed occhi piccoli, e, molto spesso, difetti cardiaci, dita soprannumerarie e sordità.
La sindrome di Edwards o trisomia 18 è caratterizzata da ritardo mentale e fisico. Gli affetti manifestano cranio allungato, orecchie con basso punto di attacco, cavità orale piccola e difetti cardiaci.
Aneuploidie cromosomi sessuali
Le anomalie più frequenti dei cromosomi sessuali sono le trisomie XXX, XXY, XYY e la monosomia X0, dove 0 sta per assenza dell’omologo del cromosoma X. Essendo la Y mascolinizzante, la sua presenza indica il sesso maschile.
La trisomia XXY o sindrome di Klinefelter è causata dalla non disgiunzione delle X nella produzione dell’ovulo. Gli individui affetti sono maschi con testicoli non sviluppati, mancata produzione di spermatozoi, assenza di pelosità e barba, tono della voce che resta acuto. In genere, tali sintomi si accompagnano a statura alta e funzione intellettuale leggermente deficitaria.
La monosomia X0 o sindrome di Turner produce femmine di bassa statura, collo tozzo e corto, organi sessuali infantili, assenza spesso di ovaie. Le capacità intellettive, in genere, risultano quasi normali.
Gli affetti della trisomia XXX sono femmine quasi normali sul piano fisico e riproduttivo, ma manifestano, molto spesso, consistenti ritardi mentali.
Gli affetti della trisomia XYY sono maschi che, a parte la tendenza a superare mediamente i 180 cm di statura, non manifestano sintomi fisici particolari. La trisomia XYY, data l’alta frequenza riscontrata nelle carceri, fu sospettata, dopo gli anni 60, di essere una probabile causa di comportamenti violenti. Studi approfonditi, però, hanno fatto rientrare del tutto tali sospetti. La trisomia XXY è causata da una mancata disgiunzione dei due cromatidi che formano il cromosoma Y durante la seconda divisione meiotica.
Malattie genetiche
Le malattie genetiche sono causate da alterazioni nel DNA di un individuo. Possono essere acquisite, come avviene nella maggior parte dei tumori, o ereditarie se l’alterazione del DNA viene trasmessa alle generazioni successive. Si classificano in
- Malattie geniche (o monogeniche o mendeliane)
- Malattie multifattoriali (o complesse o poligeniche)
- Malattie cromosomiche
- Malattie mitocondriali
Delle malattie genetiche di tipo cromosomico (aneuploidie) abbiamo già parlato.
Le malattie monogeniche o mendeliane sono causate dall’alterazione di un unico gene e seguono quindi in modo caratteristiche le leggi della ereditarietà mendeliana. La maggior parte di esse è portata da alleli recessivi. Una malattia genetica portata da alleli dominanti ha infatti molte plù probabilità di essere eliminata dalla selezione naturale, poiché si esprime sempre, sia in forma omozigote che in forma eterozigote. Le malattie genetiche portate da alleli recessivi si esprimono invece solo in forma omozigote, mentre rimangono latenti in forma eterozigote (portatori sani). Possono così ‘nascondersi’ alla selezione naturale e sopravvivere nella popolazione.
Le malattie genetiche di tipo mendeliano si classificano in:
- Malattie autosomiche recessive
- Malattie autosomiche dominanti
- Malattie X-linked (legate al cromosoma X)
- Malattie Y-linked (legate al cromosoma Y)
Malattie geniche autosomiche
Le malattie geniche autosomiche recessive sono causate da alleli recessivi presenti negli autosomi, una delle 22 coppie di cromosomi non sessuali. Come abbiamo detto, essendo portate da alleli recessivi, sono le malattie genetiche di gran lunga più diffuse e frequenti. La malattia si manifesta solo se sono presenti entrambi gli alleli e quindi gli individui malati sono necessariamente omozigoti recessivi. Gli eterozigoti sono portatori sani.
Possono essere affetti sia i maschi che le femmine.
Gli affetti sono in genere presenti in un'unica generazione (eredità orizzontale) e ricevono la malattia da entrambi i genitori portatori sani.
I genitori portatori sani hanno il 25% di probabilità di avere figli malati ad ogni gravidanza.
Per questo, per malattie ereditarie diffuse in zone circoscritte (come l’anemia mediterranea), è importante fare test genetici per individuare eventuali portatori sani i quali, se dovessero sposarsi tra loro, avrebbero una possibilità su quattro di generare figli malati.
Il modello dell’ereditarietà autosomica recessiva dimostra i principi della segregazione descritti da Mendel. Ognuno dei genitori porta un allele normale e uno difettoso e ha una probabilità del 50% al momento della meiosi di produrre un gamete con un allele difettoso. Ne consegue che la progenie avrà genotipi nella proporzione 1:2:1 e fenotipi nella frequenza di 3:1. Vale a dire che il 75% della progenie apparirà normale e il 25% evidenzierà un fenotipo recessivo (malato). La progenie "normale" avrà un rischio pari a due terzi di essere portatrice dell’allele malato (portatori sani).

Alcuni esempi di malattie autosomiche recessive sono la fenilchetonuria, la fibrosi cistica (o mucoviscidosi), l’anemia falciforme (o drepanocitosi), la beta talassemia, l’albinismo.
La fibrosi cistica (CF) è una malattia genetica caratterizzata dall’infezione cronica delle vie aeree ed è una malattia autosomica recessiva conseguente a mutazioni in un gene localizzato sul cromosoma 7.
La fenilchetonuria, detta anche PKU, è una malattia caratterizzata da un’aumentata concentrazione nel sangue e nell’urina dell’amminoacido fenilalanina e dei suoi derivati e da grave ritardo mentale.
Nell'anemia falciforme (o drepanocitosi), l'allele difettoso è codominante con quello normale: una persona eterozigote produce una certa quantità di globuli rossi a forma di falce mentre la forma normale di queste cellule del sangue è quella discoidale Questa malattia è dovuta ad un errore durante la sintesi dell'emoglobina che determina un maggiore fragilità del globulo rosso che va incontro più facilmente a lisi (rottura) con conseguenti fuoriuscite dell'emoglobina: l'individuo affetto dalla malattia presenta uno stato di anemia più o meno grave. È stata osservata una maggior frequenza della malattia in certe aree geografiche (Africa, Grecia, Sicilia e parti del Medio Oriente).
La talassemia major (o Morbo di Cooley) è una grave forma d’anemia ereditaria diffusa in tutto il bacino del Mediterraneo. I malati devono essere sottoposti a periodiche trasfusioni.
Le malattie geniche autosomiche dominanti sono causate da alleli dominanti presenti negli autosomi, una delle 22 coppie di cromosomi non sessuali.
Possono essere affetti sia i maschi che le femmine. Sono affetti gli eterozigoti, che portano l’allele malato solo in uno dei due cromosomi ereditati dai genitori. Sono estremamente improbabili individui malati omozigoti dominanti, poiché dovrebbero essere figli di genitori entrambi malati.
Gli affetti possono essere presenti in tutte le generazioni (eredità verticale). Ogni affetto ha sempre un genitore affetto (esistono però eccezioni). Ogni affetto ha il 50% di probabilità di avere figli malati ad ogni gravidanza.

Le malattie geniche autosomiche dominanti sono molto rare poiché, non potendosi mascherare nei portatori sani, sono state quasi tutte eliminate dalla selezione naturale. Tuttavia vi sono dei casi in cui le anomalie geniche dominanti possono trasmettersi ed avere una certa diffusione, in particolare quando le manifestazioni della malattia si presentano in età adulta. È questo il caso della acondroplasia, una forma di nanismo, della corèa di Huntington (ballo di SanVito) che causa una progressiva degenerazione del sistema nervoso e della distrofia muscolare miotonica. Altre affezioni di questo tipo sono: la sindrome di Marfan, la malattia di von Willebrand, la stenosi subaortica ipertrofica e la porfiria acuta.
Malattie geniche legate al sesso (sex-linked)
Malattie geniche legate al cromosoma X (X-linked)
Abbiamo già avuto modo di parlare di queste anomalie geniche nel capitolo relativo all’eredità legata al sesso (sex-linked). Si tratta di malattie portate da alleli recessivi o dominanti localizzati solo sul cromosoma X. Il cromosoma Y è più piccolo del cromosoma X e non porta l’allele corrispondente.
Nelle malattie ereditarie X-linked i geni anomali sono localizzati sul cromosoma sessuale X e di conseguenza il rischio clinico e la gravità della malattia sono diversi nei due sessi. Dato che il maschio presenta un solo cromosoma X, la presenza di un gene mutante dà luogo inevitabilmente alla manifestazione clinica morbosa, indipendentemente dall’espressività (recessiva o dominante) del carattere. Le malattie ereditarie legate al sesso non possono essere trasmesse da maschio a maschio, cioè dal padre al figlio, mentre il padre le trasmette a tutte le figlie. Nell’albero genealogico della famiglia la distribuzione delle malattie legate al sesso è diversa a seconda che si tratti di caratteri recessivi o dominanti (nella donna).
Nel caso si tratti di anomalie recessive , la malattia colpisce praticamente solo i maschi nati da madri portatrici (clinicamente sane).

Le malattie X-linked dominanti sono rare. In questo caso una madre affetta ha un 50% di probabilità d trasmettere la malattia ai propri figli (maschi o femmine che siano), mentre un padre affetto trasmette la malattia solo alle figlie.

Le principali forme ereditarie legate al sesso di tipo recessivo sono l’emofilia A, la distrofia muscolare tipo Duchenne, il deficit di glucosio-6-fosfato-deidrogenasi, il daltonismo, la sindrome dell’X-fragile. Tra le forme morbose legate al sesso dominanti hanno una certa importanza clinica lo pseudoipoparatiroidismo e il rachitismo resistente alla vitamina D.
Le malattie geniche Y-linked sono molto rare. Ne sono affetti solo i maschi che trasmettono la malattia solo ai figli maschi.

Malattie genetiche multifattoriali
In questo gruppo vengono compresi numerosi quadri morbosi (solitamente di carattere cronico-degenerativo, a carico degli adulti), quali l’ipertensione essenziale, la psoriasi, l’epilessia, le malattie coronariche, il diabete mellito, l’ulcera peptica, l’asma bronchiale, alcune forme di depressione che presentano un caratteristico andamento familiare e i cui meccanismi patogenetici comprendono una serie di geni (più o meno alterati) che interagiscono in maniera cumulativa fino a dare luogo alla manifestazione morbosa. In altri termini, la componente ereditaria di queste affezioni si manifesta nell’interazione di molteplici fattori „predisponenti“ (su base genetica) con fattori multipli ambientali. Alcune persone possiedono per esempio una predisposizione genetica al cancro al polmone, ma potranno evitarlo evitando di fumare. Altri individui con predisposizione al diabete potranno non ammalarsi se terranno sotto controllo il peso corporeo. Dato che il numero esatto dei geni responsabili di questi tratti poligenici non è noto, è assai difficile calcolare con precisione il rischio che un soggetto presenta di ereditare una certa condizione morbosa.
Malattie genetiche mitocondriali
I mitocondri sono dotati di un proprio DNA (mtDNA). L’mtDNA è estremamente piccolo, ha struttura circolare, e contiene solamente 37 geni: 13 geni che codificano alcune delle subunità proteiche dei complessi della catena respiratoria e 24 geni che codificano molecole indispensabili alla sintesi di tali subunità (2 RNA ribosomali, rRNA e 22 RNA transfer, tRNA).
Durante la fecondazione, i mitocondri presenti nel nuovo individuo (zigote) provengono solo dalla cellula uovo. Una madre portatrice di una mutazione del mtDNA trasmetterà tale mutazione a tutti i propri figli, ma solo le figlie trasmetteranno a loro volta la mutazione alla loro progenie (eredità matrilineare).
La principale funzione dei mitocondri è quella di produrre energia. Quando questa energia viene a mancare, per un alterato funzionamento mitocondriale, la vita della cellula stessa e, conseguentemente, quella del nostro organismo sono a rischio. I mitocondri sono presenti in tutti i tessuti pertanto, le malattie mitocondriali, possono colpire qualsiasi organo, ma i più colpiti sono i tessuti e gli organi che “consumano” più energia: cervello, muscoli, cuore. Tali malattie, nelle diverse varianti, sono accomunate dall’induzione di un deficit energetico e sono comunemente definite encefalomiopatie mitocondriale. Dal punto di vista clinico sono molto variabili per quanto concerne età di esordio, tipo di evoluzione e tessuto coinvolto.
Nei neonati, tali malattie, possono manifestarsi con gravi anemie e disfunzioni pancreatiche comunemente note come sindrome del bambino flaccido (floppy infant) o coma cheto-acidotico con disfunzione epatorenale. Nell’infanzia possono rallentare o arrestare la crescita, causare danni renali, nanismo, disfunzioni endocrine (diabete mellito, diabete insipido), atrofia ottica, sordità, encefalopatia progressiva. Nell’età adulta, i sintomi ricorrenti, sono intolleranza allo sforzo ed affaticamento.
Ingegneria Genetica
L’ingegneria genetica (più propriamente tecnologie del DNA ricombinante) è un insieme di tecniche (biotecnologia) che ha l’obiettivo di inserire, eliminare, inattivare o modificare geni all’interno di un organismo, producendo organismi geneticamente modificati (OGM).
Il termine ‘OGM’ è improprio da un punto di vista biologico in quanto tutti gli organismi caratterizzati da riproduzione sessuata sono, a rigor di termini, geneticamente modificati (nessun individuo è uguale ai propri genitori e nemmeno ai propri fratelli).
La definizione adottata dalla Direttiva europea 2001/18, che regola il rilascio ambientale degli OGM, è la seguente: «un organismo, il cui materiale genetico è stato modificato in modo diverso da quanto avviene in natura con l’accoppiamento e/o la ricombinazione genica naturale». Tale modifica viene definita con il termine di “transgenesi” e l’organismo da esso derivato viene detto transgenico o ricombinante.
Tutto ciò che viene invece ottenuto con programmi di miglioramento genetico convenzionale (incrocio e selezione) «inclusa la mutagenesi (mutazioni indotte dall'esposizione a radiazioni o ad mutageni chimici.) e la fusione cellulare di cellule vegetali di organismi che possono scambiare materiale genetico anche con metodi di riproduzione tradizionali» è escluso dalla definizione di OGM, pur comportando modificazioni del genoma spesso di gran lunga superiori. Tali organismi mutati, non essendo classificati come OGM sono ammessi all'uso anche in agricoltura biologica e molte di queste cultivar (varietà vegetali) sono attualmente presenti sul mercato ed apprezzate dai consumatori. Il popolare grano duro italiano (varietà Creso), ad esempio è una mutazione ottenuta nel 1974 irraggiando la varietà “Cappelli” con raggi gamma provenienti da scorie di reattori nucleari.
Si tenga quindi presente che modificazioni del patrimonio genetico di piante ed animali erano state ottenute anche precedentemente l’avvento delle tecniche di ingegneria genetica. Ma con il termine OGM si intende fare una distinzione tra gli organismi il cui patrimonio genetico è stato modificato tramite l'uso di tecniche di miglioramento genetico classico, considerate "naturali", da quelli modificati tramite la tecnica del DNA ricombinante, che consentono l'inserimento mirato di sequenze geniche che conducono al fenotipo desiderato.
La maggior parte dei risultati finora conseguiti attraverso l’utilizzo delle tecniche de DNA ricombinante sono stati ottenuti attraverso l’isolamento di geni e la successiva introduzione in un ospite eterologo, anche appartenente ad una specie differente (una sorta di trapianto genico), al fine di conferire caratteristiche nuove alle cellule riceventi. L’obiettivo è quello di esprimere una proteina nuova all'interno di un organismo diverso da quello di origine allo scopo di conferirgli caratteristiche utili.
Attualmente le applicazioni più promettenti si hanno in campo agroalimentare, biomedico ed ambientale.
Applicazioni agroalimentari
- piante resistenti agli erbicidi
- piante resistenti ai parassiti
- piante resistenti alla siccità
- frutti che si conservano più a lungo
- alimenti arricchiti dal punto di vista nutrizionale (vitamina A nel riso, acidi grassi nella soia).
Applicazioni biomediche ed ambientali
- Sintesi di molecole (microrganismi in grado di sintetizzare farmaci, ormoni, vaccini, enzimi, antibiotici etc)
- depurazione (batteri che degradano idrocarburi, piante capaci di stoccare metalli pesanti)
- Terapia genica (sostituzione geni alterati, introduzione di geni in cellule malate che blocchino il malfunzionamento, introduzioni di geni in cellule tumorali che ne inducano la morte)
- test DNA o DNA fingerprinting (letteralmente impronta digitale del DNA, individuazione e confronto DNA per test di paternità, casi di omicidio)
Le tecnologie del DNA ricombinante si avvalgono di tre strumenti molecolari fondamentali:
- i vettori genici particolari molecole di DNA in grado di ospitare un gene estraneo e di introdurlo in un genoma.
- gli enzimi di restrizione in grado di ‘ritagliare’ un frammento di DNA contenente il gene e di ‘aprire’ i vettori genici in modo tale che entrambi, vettore ed inserto genico, presentino le estremità complementari e facilmente ricomponibili
- le DNA-ligasi, enzimi in grado di ricomporre i frammenti in modo tale che il gene si integri nel vettore genico.
Gli enzimi di restrizione
Gli enzimi di restrizione sono delle endonucleasi, enzimi della famiglia delle nucleasi, che i batteri utilizzano per difendersi dai virus che li infettano, facendo a pezzi il DNA virale. Le nucleasi si distinguono in endonucleasi, che tagliano il filamento di DNA in un punto interno dividendolo in due frammenti ed esonucleasi, che degradano il DNA asportandone nucleotidi alle estremità.
Gli enzimi di restrizione presentano la notevole peculiarità di tagliare il DNA solo in corrispondenza di particolari sequenze di basi (in genere 4, 6 o 8 basi) che essi riconoscono (siti di restrizione).
I siti di restrizione sono quasi tutti palindromi, cioè la successione delle loro basi presenta una sequenza ad essa complementare che risulta uguale se letta al contrario. Ad esempio la successione ACTAGT ha come complementare la successione TGATCA (che è uguale alla precedente letta al contrario). Per convenzione le sequenze di riconoscimento sono scritte da sinistra a destra in direzione 5’ → 3’. Quindi ACTAGT rappresenta la sequenza 5’- ACTAGT -3’.
(Una frase è palindrome quando è uguale se letta nei due sensi: “i topi non avevano nipoti”)
In un filamento di DNA si trovano naturalmente numerosi siti di riconoscimento per una particolare endonucleasi. In questo modo le endonucleasi tagliano il DNA in molti segmenti, detti frammenti di restrizione.
Gli enzimi di restrizione possono tagliare il DNA in due modi. Alcuni enzimi tagliano il DNA lasciandone le estremità piatte, altri lasciando le estremità sfalsate (alcuni con il filamento 5’ più lungo del filamento 3’, altri viceversa).
Gli enzimi di restrizione che generano estremità sfalsate sono quelli comunemente usati in ingegneria genetica. Infatti tutte le estremità generate dagli enzimi che operano un taglio sfalsato, siano esse sporgenti in 5’ o in 3’, sono “coesive” (o adesive o appiccicose, sticky ends), cioè possono formare ponti idrogeno tra le due code a filamento singolo complementari. Nel caso l’enzima di restrizione produca estremità piatte è possibile aggiungere una “coda” (tailing) sfalsata, usando l’enzima terminal transferasi, che aggiunge nucleotidi alle estremità 3’ del DNA, generando estremità coesive.
Le estremità coesive facilitano la reazione di ricomposizione della DNA ligasi.
In questo modo, mescolando frammenti di restrizione, anche provenienti da DNA diverso, tagliati con la medesima endonucleasi, i frammenti aderiscono spontaneamente con le loro estremità coesive per un tempo sufficientemente lungo da rendere efficace l’azione di saldatura (formazione dei legami fosfo-diestere) della DNA-ligasi.
L’enzima di restrizione EcoRI (leggi “Eco erre primo”), ad esempio, si lega a sequenze specifiche di DNA in doppia elica GAATTC. Taglia (idrolizza) in modo sfalsato i legami fosfodiestere tra Guanina ed Adenina su entrambi i filamenti (il taglio produce gruppi 3’-OH e 5’-fosfato), generando frammenti di DNA che hanno quattro basi sporgenti in singolo filamento alle estremità 5’ (estremità 5’ sporgenti).

La nomenclatura degli enzimi di restrizione prevede:
- Le prime tre lettere, scritte in corsivo, sono prese dalla nomenclatura binomiale del batterio di origine.
- Sierotipi differenti dello stesso organismo sono identificati da una quarta lettera minuscola (Es. Hind, Hinf).
- Segue una lettera maiuscola o un numero, che identificano un ceppo particolare di quel batterio, ove fosse necessario.
- Un numero romano indica l’ordine di scoperta, qualora dallo stesso batterio vengano isolati enzimi diversi.
Ad esempio:
Pst I Providencia stuarti (I enzima isolato)
Eco RI Escherichia coli Ry13 (I enzima isolato)
Eco RII Escherichia coli R245 (II enzima isolato)
cDNA (DNA complementare)
Gli enzimi di restrizione non sono l’unico modo per ottenere un inserto genico da inserire in un vettore. Attraverso l’enzima trascrittasi inversa (o retrotrascrittasi) è possibile catalizzare la trascrizione dell’mRNA in DNA. Dopo che è avvenuta la sintesi di un filamento singolo di DNA si assembla il secondo filamento di DNA, usando il primo come stampo. Il DNA che si forma in questo modo è detto cDNA (DNA complementare). Questa tecnica è utile quando si vuole far esprimere un gene umano ad un batterio. I geni umani sono infatti costituiti di sequenze codificanti (esoni) e sequenze non codificanti (introni) ed i batteri non possiedono le strutture adatte per eliminare gli introni prima di sintetizzare la proteina corrispondente. La molecola di mRNA che si è formata a partire dal gene ha già eliminato gli introni (splicing). Ricostruendo quindi il gene a partire dall’mRNA si ottiene una sequenza genica ‘pulita’ che i batteri potranno poi trasformare nella proteina corrispondente.
Vettori genici
I vettori genici sono delle sequenze di DNA di diversa natura nelle quali sia possibile inserire delle altre sequenze nucleotidiche (gene esogeno) e che siano in grado di integrarsi con il DNA-bersaglio introducendovi il gene esogeno. Attualmente sono disponibili diversi tipi di vettori genici, la cui scelta dipende dalla natura (animale, vegetale, procariote) della cellula bersaglio che deve ricombinare il suo DNA e dalle dimensioni dell’inserto genico che devono ospitare.
VETTORI GENICI |
DIMENSIONI DELL’INSERTO |
Plasmide |
0 -10 kbp |
Fago λ |
10 -20 kbp |
Cosmide |
30 -45 kbp |
Fago P1 |
70 – 100 kbp |
PAC (cromosomi artificiali P1 ) |
130 – 150 Kbp |
BAC (cromosomi batterici artificiali) |
0 – 300 Kbp |
YAC (cromosomi artificiali di lievito) |
0,2 – 2 Mbp |
MAC (cromosomi artificiali di mammifero) |
> 1 Mbp |
I vettori più usati per i batteri (E.coli) sono i plasmidi e i derivati da virus batteriofagi.
Nel caso di vegetali si utilizza un batterio parassita endocellulare di molte specie di piante, l’Agrobacterium tumefaciens, il cui plasmide Ti è in grado di inserirsi nel genoma della cellula vegetale.
Per il lievito (Saccharomyces cerevisiae), un eucariote unicellulare, si utilizza il suo plasmide naturale 2 μm (lunghezza della sua circonferenza) ed una serie di vettori ingegnerizzati da esso derivati.
Infine, per ricombinare cellule animali si utilizzano prevalentemente vettori di natura virale.
CELLULA OSPITE |
VETTORI GENICI COMPATIBILI |
Batterio |
Plasmidi, fago λ, fago M13, fago P1, PAC, BAC, YAC |
Lievito |
Plasmide 2 μm, Plasmidi “navetta” (shuttle), vettori ARS, YAC |
Cellule animali |
Vettori derivati da virus |
Cellule vegetali |
Plasmidi derivati dal batterio Agrobacterium tumefaciens |
Caratteristiche di un vettore genico
In generale, per svolgere la sua funzione, un vettore deve possedere le seguenti proprietà:
• Un segmento di DNA che lo renda capace di replicarsi nella cellula ospite (replicon) o di integrarsi al suo genoma. Il DNA del replicone interagisce con gli enzimi coinvolti nell’avvio della replicazione del DNA. Un replicone è un’unità genetica composta da un’origine di replicazione del DNA (ori) e dagli elementi di controllo ad essa associati. Si può anche definire come il più piccolo pezzo di DNA in grado di replicarsi nell’ospite mantenendo un numero stabile di copie. Nei plasmidi, un “replicone” è lungo in media 500 bp.
• Uno o più marcatori genetici, la cui presenza permetta di selezionare le cellule che ospitano il plasmide. I marcatori genetici possono essere resistenze ad antibiotici o marcatori nutrizionali.
• Singoli siti di taglio per il più alto numero possibile di enzimi di restrizione. I vettori sintetici (ingegnerizzati) più recenti presentano un unica regione (sito di policlonaggio o polylinker) che riunisce i siti di riconoscimento per gli enzimi di restrizione più comuni. Questi siti devono stare al di fuori delle regioni essenziali (replicone e marcatore).
Non è essenziale, ma può essere molto utile inserire nel vettore degli elementi di controllo dell’espressione genica. Uno dei più usati è l’operone lac che in E.coli controlla la sintesi degli enzimi necessari all’utilizzo del lattosio da parte del batterio. L’operone è formato da più geni che interagiscono. Uno di questi, il promotore Plac, attiva i geni a valle in presenza di lattosio. È dunque sufficiente inserire il gene di interesse a valle del promotore Plac per avere a disposizione un meccanismo semplice di controllo della sua espressione. In presenza di lattosio (o di molecole analoghe che non possono essere metabolizzate da E. coli) la cellula sintetizza la proteina codificata dal gene che ci interessa, mentre in assenza di lattosio il gene di interesse non si esprime. Vettori che contengono regioni per il controllo dell’espressione sono detti vettori d’espressione.
L’operone lac è anche un buon esempio di marcatore nutrizionale utilizzabile, assieme ad una resistenza ad un antibiotico, per selezionare le cellule che ospitano il plasmide ricombinante. L’operone lac contiene infatti la sequenza-bersaglio dell’enzima di restrizione e quindi se il plasmide riceve l’inserto del DNA donatore, questo si inserisce all’interno dell’operone e la funzione di digestione del lattosio viene persa.
Le cellule trasformate dal plasmide ricombinato acquistano la resistenza all’antibiotico, ma non la capacità di digerire il lattosio (infatti il gene LacZ viene interrotto dall’inserzione del DNA fagico),
le cellule che si trasformano col plasmide non ricombinato acquistano entrambe le proprietà,
le cellule che non acquistano alcun plasmide restano sensibili all’antibiotico.
Plasmidi
I primi vettori genici individuati ed utilizzati sono stati i plasmidi.
I plasmidi sono brevi sequenze anulari di DNA batterico extracromosomiche. Tutti i batteri possiedono un unico filamento di DNA chiuso ad anello, detto cromosoma batterico. Alcuni batteri possiedono inoltre alcuni filamenti anulari più piccoli, contenenti qualche decina di geni, detti plasmidi, che sono in grado di duplicarsi indipendentemente dal cromosoma batterico.
Se sottoposti a stress ambientale (carenza alimentare, antibiotici) i batteri effettuano un processo di coniugazione, attraverso il quale una cellula donatrice (F+) trasferisce copia di uno o più plasmidi ad una cellula ricevente (F-). Durante il processo di coniugazione due batteri si uniscono attraverso un ponte citoplasmatico (pilo sessuale). Nella cellula donatrice il plasmide inizia a replicarsi a partire da un punto, detto ‘sito di origine della replicazione’ (ori). Il filamento di DNA che viene sostituito dal nuovo filamento penetra attraverso il pilo nella cellula ricevente.
La cellula ricevente duplica a sua volta il monofilamento ricevuto rigenerando il plasmide completo. I geni trasferiti sono successivamente integrati nel cromosoma batterico.

I batteri si riproducono asessualmente per mitosi. Ciò significa che gli individui di ogni nuova generazione sono geneticamente identici a quelli della generazione precedente. Questa uniformità genetica è evidentemente controproducente. La probabilità di adattarsi alle variazioni ambientali è tanto più elevata quanto maggiore è la variabilità genetica all’interno di una popolazione. I batteri risolvono questo problema tramite la coniugazione, un fenomeno attraverso il quale ricombinano il loro patrimonio genetico, creando nuova variabilità ed aumentando le probabilità di sopravvivenza.
La coniugazione batterica è un fenomeno di sessualità.
Da un punto di vista biologico infatti riproduzione e sessualità sono fenomeni diversi. La riproduzione genera nuovi individui. La sessualità genera variabilità genetica. Negli organismi superiori, dove avviene una riproduzione sessuata, i due processi avvengono contemporaneamente. La formazione dei gameti necessari alla riproduzione avviene infatti attraverso processi sessuali di rimescolamento dei caratteri genetici (crossino-over, assortimento indipendente degli omologhi) che garantisce una elevata variabilità genetica.
Per questa loro capacità di entrare in un batterio ed integrarsi con il cromosoma batterico, i plasmidi sono dei vettori ideali per inserire geni esogeni e farli esprimere ai batteri. È in questo modo, ad esempio, che per la prima volta si è riusciti a far sintetizzare a Escherichia coli insulina umana.
Tra i diversi tipi di plasmidi che i batteri sono in grado di trasferire, sono particolarmente importanti ed utilizzati nelle tecniche del DNA ricombinante i plasmidi R, che contengono geni che conferiscono al batterio la resistenza agli antibiotici. Questa caratteristica è utilizzabile per discriminare facilmente i batteri che hanno ricevuto il plasmide con il gene esogeno da quelli che non l’hanno ricevuto. È infatti sufficiente trattare la popolazione batterica con l’antibiotico di cui il plasmide porta il fattore di resistenza: solo i batteri ricombinanti ovviamente sopravviveranno. Attualmente si utilizzano plasmidi che portano geni per la resistenza alla ampicillina (amp) o alla canamicina (kan).
Per essere utilizzato come vettore genico un plasmide deve poi possedere un unico sito di attacco per l’enzima di restrizione che si vuole usare. In questo modo, trattato con l’enzima di restrizione, il plasmide si aprirà in un solo punto, dove poi si salderà il gene esogeno.

Tra tutti i vari vettori disponibili, i plasmidi hanno acquistato un posto privilegiato per la loro affidabilità e facilità di manipolazione. Semplicemente, lavorano meglio degli altri vettori se il DNA da clonare non è di grandi dimensioni

In sintesi dunque le fasi per la produzione di un plasmide ricombinante sono:
- Il DNA sorgente (o donatore) viene sottoposto ad enzimi di restrizione che lo tagliano in frammenti, uno dei quali conterrà il DNA bersaglio (inserto) con il gene di interesse.
- Gli stessi enzimi vengono poi usati per creare sul plasmide un punto di rottura complementare in cui poter inserire il DNA esogeno.
- Mediante l'azione della DNA-ligasi si saldano i frammenti del DNA sorgente al plasmide.

È possibile anche utilizzare plasmidi con due siti di riconoscimento per enzimi di restrizione diversi ed utilizzare gli stessi due enzimi per tagliare il frammento di DNA bersaglio da inserire. Si ottengono frammenti con estremità coesive differenti. Il vantaggio di questa strategia è che, siccome solo le estremità “sorelle” sono compatibili tra loro, l’inserto di DNA si lega al vettore in un solo modo (clonazione direzionale), permettendo di conoscere in anticipo l’orientamento dell’inserto. Inoltre il vettore tagliato con due enzimi diversi non circolarizza (non si richiude prima di aver saldato l’inserto), perché le due estremità non sono compatibili.
Il plasmide ricombinante a questo punto è pronto per essere introdotto nelle cellule batteriche. Il trasferimento di un plasmide ricombinante avviene sfruttando in genere il fenomeno della trasformazione batterica.
La trasformazione batterica è un processo naturale, attraverso il quale alcuni procarioti (detti competenti) sono in grado di ricevere del DNA esterno in grado di produrre nuove caratteristiche. Questo fenomeno fu scoperto nel 1928 da Frederick Griffith. Prima dell’avvento degli antibiotici, i pazienti affetti da polmonite venivano trattati con antisieri preparati iniettando cellule morte del batterio agente della malattia (Streptococcus pneumoniae). Il medico inglese Frederick Griffith nel tentativo di preparare antisieri più efficaci si imbatté in risultati inattesi. Streptococcus pneumoniae è un batterio la cui parete è rivestita da una spessa capsula. Gli zuccheri della capsula provocano la risposta immunitaria dell’ospite. Un sierotipo è un ceppo batterico con proprietà immunologiche particolari, che gli sono conferite dalla mistura di zuccheri della capsula. Griffith dimostrò che i batteri avirulenti, a contatto con i batteri virulenti morti, ne acquisivano sempre il sierotipo. Il processo di conversione dei batteri avirulenti innocui in cellule virulenti fu chiamato trasformazione. Gli esperimenti di Griffith hanno un’importanza storica notevole, perché portarono Osvald Avery (Columbia University, New York) ad indagare quale fosse il principio trasformante, che permetteva ai batteri avirulenti di acquistare le proprietà delle cellule morte. Nel 1944 Avery, MacLeod e McCarty dimostrarono che il principio trasformante era il DNA. La cellula ricevente assorbe dall’ambiente un frammento “nudo” di DNA rilasciato da una cellula donatrice. Tuttavia la rigida parete cellulare dei batteri è di grande ostacolo al passaggio delle macromolecole di DNA. Pochi generi di batteri (Streptococcus, Neisseria, Hemophilus) presentano lo stato di competenza alla trasformazione in modo naturale durante una fase del loro ciclo vitale, in cui riformano la parete cellulare. Questi batteri possiedono una competenza naturale alla trasformazione e non richiedono nessun trattamento particolare per indurre la loro capacità di assumere il DNA. Virtualmente, tutti i batteri hanno la capacità di assumere DNA “nudo” dall’ambiente, posto che acquistino una “competenza” alla trasformazione. La maggioranza dei batteri non è naturalmente competente alla trasformazione, per cui è necessario indurre uno stato di competenza artificiale. Oggi sono state sviluppate alcune tecniche chimiche e fisiche, per quanto molto empiriche, in grado di rendere competenti anche batteri che non lo sono naturalmente. I batteri gram-negativi diventano competenti se trattati con agenti chimici o fisici. I batteri gram-positivi non possiedono la membrana esterna propria degli organismi gram-negativi. Tuttavia l’ingresso del DNA è impedito dalla presenza di uno strato rigido esterno di mureina o peptidoglicani, che è molto più spesso del suo equivalente nei batteri gram-negativi. Questo strato deve essere preventivamente idrolizzato con lisozima, quindi i protoplasti (cioè i batteri senza parete cellulare) si possono trasformare.
- I metodi chimici si basano sull’osservazione che batteri trattati con soluzioni fredde di cationi bivalenti lasciano entrare DNA nudo in modo più efficiente. Si ipotizza che l’aggiunta di ioni bivalenti (Ca2+), mascherando le cariche negative del DNA, ne favorisca l’ingresso nella cellula. L’introduzione è favorita da brevi periodi di shock termico.
- I metodi fisici si basano sull’osservazione che la presenza di un campo elettrico destabilizza le membrane battericheed induce la formazione di pori transienti del diametro di alcuni manometri (elettroporazione). Le molecole di DNA passano attraverso questi pori applicando impulsi di voltaggio elevato (12,5-15 kV/cm)..
Un limite all’utilizzo del plasmide come vettore è dato dalla dimensione del frammento del DNA che esso può effettivamente contenere ed al fatto che non può essere utilizzato per trasformare cellule animali. Sono stati quindi sviluppati altri sistemi vettore.
Vettori virali
Un’importante famiglia di vettori genici è costituita dai vettori virali. I virus sono costituiti da un involucro di proteine (capside) di ridottissime dimensioni al cui interno è racchiuso il DNA virale contenente solo le informazioni essenziali e quindi non possono riprodursi da soli. Per farlo devono penetrare all’interno di una cellula ed utilizzarne le strutture di sintesi. Quando un virus infetta una cellula, integra il suo DNA con il genoma cellulare. In questo modo la cellula esprime le informazioni virali e sintetizza centinaia di copie del virus. Durante questo processo possono avvenire scambi tra il DNA virale ed il DNA cellulare. In laboratorio è possibile pilotare questo fenomeno di ricombinazione tra i due genomi, al fine di trasferire geni particolarmente interessanti nelle cellule. Per far ciò, si sostituiscono alcuni geni del virus con geni utili, di qualunque provenienza, ed un gene marcatore per verificare l'avvenuto trasferimento. Il virus modificato è ancora in grado di infettare la cellula, ma, anziché sfruttarla e distruggerla, la arricchisce delle nuove caratteristiche desiderate. Ogni virus utilizza le proteine del suo capside per riconoscere ed infettare un particolare tipo di cellula.
I vettori virali sono utilizzati soprattutto per introdurre geni esogeni nei batteri (in alternativa ai plasmidi) e nelle cellule eucarioti animali.
I virus che infettano i batteri sono detti batteriofagi o fagi. Quando un fago infetta un batterio può acquisirne frammenti di DNA che vengono ceduti alla cellula batterica ricevente durante il successivo processo di infezione. Il trasferimento di materiale genetico da un batterio ad un altro tramite fagi è noto come trasduzione. La trasduzione è dunque una sorta di trasformazione batterica mediata da virus. I biologi molecolari sfruttano dunque questa capacità dei virus per utilizzarli come vettori genici. Alcuni fagi (Lambda ed M13) sono stati adattati alle esigenze dei biologi molecolari che ne hanno opportunamente modificato il DNA per dotarlo di specifici siti di restrizione ed hanno eliminato quella parte del genoma non indispensabile per la replicazione. Il DNA virale di un fago, una volta penetrato in una cellula batterica, può integrarsi nel cromosoma batterico e rimanere in fase latente (profago) per molto tempo senza interferire sulla vita della cellula (ciclo lisogenico o lisogeno) oppure può attivarsi ed utilizzare le strutture della cellula batterica per riprodursi e distruggere la cellula ospite (ciclo litico).
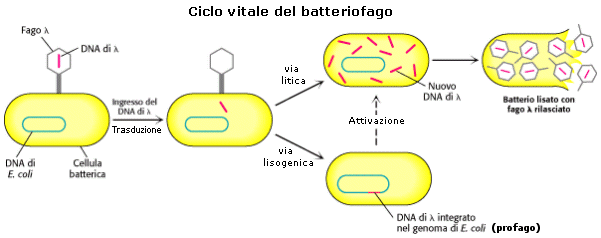
I fagi che causano la morte dei batteri per lisi cellulare sono detti virulenti (solo ciclo litico), mentre i fagi che riescono a stabilire un rapporto di simbiosi con l’ospite si dicono temperati (alternanza ciclo litico e lisogenico), come ad esempio il fago λ, ampiamente utilizzato come vettore genico. Vi sono due modalità di trasduzione. Nel corso del ciclo litico di molti virus il DNA della cellula ospite si frammenta e quando vengono assemblate le nuove particelle virali, alcuni di questi frammenti di DNA, spezzati a caso, possono essere erroneamente racchiusi all’interno del capside al posto del DNA virale (trasduzione generalizzata).

Nel corso del ciclo lisogenico il profago può staccarsi dal cromosoma batterico in cui si era integrato (escissione) portandosi via un frammento di DNA batterico ad esso adiacente (trasduzione specializzata).
 Il fago λ è utilizzato per clonare frammenti di DNA lunghi fino a 25 kbp. Normalmente il genoma del fago è lungo 49 kbp ma viene ridotto eliminando la porzione centrale con enzimi di restrizione che lasciano estremità coesive, lo stesso enzima con cui si taglia il frammento da clonare che vi si lega. Il profago integrato nel genoma si libera in seguito a danni del DNA. Prolifera a spese dell’ospite e lisa la cellula infetta liberando circa 100 virioni
Il fago λ è utilizzato per clonare frammenti di DNA lunghi fino a 25 kbp. Normalmente il genoma del fago è lungo 49 kbp ma viene ridotto eliminando la porzione centrale con enzimi di restrizione che lasciano estremità coesive, lo stesso enzima con cui si taglia il frammento da clonare che vi si lega. Il profago integrato nel genoma si libera in seguito a danni del DNA. Prolifera a spese dell’ospite e lisa la cellula infetta liberando circa 100 virioni
Cosmidi
Il DNA del fago λ, isolato dalle particelle virali è una molecola lineare con estremità 5’-sporgenti di 12 nucleotidi, dette terminali coesivi (cos). Le estremità cos circolarizzano subito dopo l’infezione della cellula ospite, grazie alla DNA ligasi di E. coli. Oltre ad essere importanti per la sopravvivenza di λ all’interno di E. coli, le estremità cos giocano un ruolo importante per l’impacchettamento del DNA nei virioni e per lo sviluppo di vettori artificiali, i cosmidi, che possono trasportare inserti di DNA di grandi dimensioni (30-45 kbp).
I cosmidi sono ibridi tra plasmidi (replicone, resistenza ad antibiotici) e le estremità cos del fago λ. La capacità massima degli inserti (45 kbp) è limitata dalle dimensioni della testa di λ.

Il cosmide ricombinante viene poi impacchettato in particelle fagiche in vitro, posto che vengano forniti i precursori delle teste, le code e le proteine per il montaggio. Il cosmide ricombinante viene successivamente introdotto nel batterio per trasduzione.
Vettori navetta (shuttle)
I vettori navetta sono vettori ingegnerizzati compatibili con cellule diverse, ad esempio batteri/lieviti o batteri/animali. In genere è fornito di due siti di replicazione, ognuno dei quali è riconosciuto e funziona su di un ospite cellulare diverso. Un vettore di questo tipo consente ad esempio la manipolazione del vettore nelle cellule batteriche ed il successivo trasferimento nelle cellule umane dove poter effettuare le analisi di interesse.

Vettori del lievito
Il lievito Saccharomyces cerevisiae rappresenta per gli eucarioti ciò che E.coli è per i procarioti. S.cerevisiae possiede un plasmide naturale di 2 μm di circonferenza (Plasmide 2 μm) che può essere utilizzato come vettore genico. Questo plasmide è stato variamente manipolato per ottenere vettori artificiali di lievito. Ad esempio, l’aggiunta al plasmide di un centromero proveniente da un cromosoma del lievito stesso garantisce che, durante la divisione cellulare del lievito i plasmidi in esso inseriti vengano ripartiti equamente in tutte le cellule che si formano (il fuso riconosce il centromero e tratta i plasmidi come cromosomi).
Se oltre al centromero si aggiungono al plasmide, dopo averlo aperto, anche dei telomeri si ottiene un vettore chiamato Cromosoma artificiale di lievito (YAR).
Se questo vettore contiene le origini di replicazione del lievito (sequenze di replicazione autonome, ARS), durante la divisione cellulare si comporta come un piccolo cromosoma di lievito.
Cromosomi artificiali
Oggi sono stati messi a punto cromosomi artificiali anche per batteri e mammiferi che contengono gli elementi genetici utili per la stabilità e la funzionalità del cromosoma (telomeri, centromeri) . Con essi è possibile utilizzare i batteri per clonare grossi pezzi di DNA (200-500 kbp).
Sono cromosomi artificiali:
- YAC, cromosomi di lievito (Yeast Artificial Chromosomes)
- BAC, cromosomi batterici
- MAC, cromosomi di mammifero
- PAC, cromosomi P1
Piante transgeniche
Le cellule vegetali sono particolarmente resistenti alla trasformazione perché sono rivestite esternamente da una spessa parete cellulare. La principale tecnica di modificazione genetica di vegetali è legata all'attitudine naturale del batterio Agrobacterium tumefaciens di infettare piante e causare una crescita paragonabile a quella tumorale negli animali, formando una moltiplicazione incontrollata delle cellule infettate entro cui il batterio si può riprodurre. A. tumefaciens contiene un plasmide (plasmide Ti, Tumor inducing) che può essere trasferito all'interno della pianta infettata ed integrare nel genoma della stessa la regione T-DNA (DNA che induce il tumore). Questo gene produce la proliferazione cellulare, che ospita il batterio e ne fornisce nutrienti per la crescita.
I biologi molecolari sono in grado di creare una popolazione di Agrobacterium che abbia nel T-DNA, al posto dei geni che causano il tumore, il gene che si desidera trasferire nella pianta. Il batterio modificato viene poi utilizzato per infettare alcuni frammenti di tessuto vegetale. Le cellule che ricevono il plasmide T iniziano a dividersi producendo calli (masserelle di cellule indifferenziate) visibili ad occhio nudo. Frammentando il callo e trattandolo con una miscela calibrata di ormoni vegetali si rigenerano le pianticelle che possiedono il gene nuovo (transgeniche). Questo sistema rapido, semplice ed efficiente, funziona tuttavia solo con piante dicotiledoni (es. pisello, fava, soia, lenticchie, ecc.) e non con le monocotiledoni, famiglia che comprende i cereali e quindi molto importante dal punto di vista economico.
Per introdurre geni esogeni in piante monocotiledoni si utilizzano il metodo biobalistico o il metodo dei protoplasti.
Il metodo biobalistico (anche detto gene gun, pistola a geni), permette di "sparare" ad altissima velocità microproiettili di oro o tungsteno ricoperti di DNA all'interno delle cellule vegetali. Tale metodo è ampiamente usato, ad esempio per la produzione del più comune cereale OGM, il mais BT.
Con il metodo dei protoplasti le cellule vegetali sono trattate con enzimi litici (cellulasi) che sciolgono la parete cellulare, trasformando le cellule in protoplasti. Questi sono più facili da trasformare, potendo assorbire DNA nudo, soprattutto se vengono stimolati elettricamente (elettroporazione).
Animali transgenici e terapia genica
Gli animali possono essere geneticamente modificati in modo del tutto naturale attraverso le infezioni virali. La modificazione genetica, in ogni caso, avviene solo se il virus penetra nella cellula bersaglio, senza risposta del sistema immunitario ed il suo genoma penetra fino al nucleo di essa.
La proprietà dei virus di integrarsi con il genoma della cellula animale (provirus) sono sfruttate dai ricercatori per progettare vettori in grado di modificare in modo controllato il genoma delle cellule animali. In medicina, questa proprietà ha aperto la strada alla Terapia genica che consentirebbe di sostituire frammenti di genoma mutato nei pazienti, agendo proprio sulle cellule adulte che hanno subito mutazioni. Le mutazioni prodotte dalla terapia genica sono dunque totalmente a carico della linea somatica e non di quella germinale.
Selezione delle cellule transgeniche
Naturalmente durante le operazioni di transgenesi alcune cellule possono non introdurre i vettori. Bisognerà perciò eseguire una selezione che dovrà eliminare le cellule che non hanno introdotto il vettore. Sarà sufficiente trattare le cellule con l’antibiotico, il cui fattore di resistenza è presente nel vettore utilizzato. Le cellule che hanno incorporato il vettore, cresceranno nel terreno contenente l’antibiotico, mentre le altre moriranno.
Librerie genomiche
Se il DNA bersaglio è già stato selezionato tra i frammenti di restrizione prima di essere integrato nel vettore o si possiede già il filamento di cDNA, le tecniche di clonazione e selezione sono relativamente semplici e si parla di clonazione mirata.
In genere è sufficiente analizzare pochi cloni ricombinanti per individuare il clone giusto che contiene il DNA bersaglio.
Se invece viene isolato e frammentato con enzimi di restrizione l’intero genoma di una cellula ed i diversi frammenti ottenuti vengono integrati ognuno in un vettore e successivamente clonati (clonazione a raffica o shotgun), si ottiene una popolazione complessa di batteri ricombinanti, ciascuno dei quali contiene un inserto diverso di DNA.
L’insieme delle colonie batteriche contenenti ognuno un diverso frammento di DNA costituisce una libreria genomica o genoteca.

Se ora si vuole individuare all’interno della genoteca la colonia batterica che contiene un particolare frammento genico sarà necessario usare una sonda che la riconosca e ce la renda evidente. Una sonda è costituita da una breve sequenza di una ventina di basi complementare al DNA di interesse, marcata mediante l’impiego di isotopi radioattivi o di un composto fluorescente. Una sonda viene in genere costruita tramite trascrizione inversa a partire dall’mRNA con formazione di cDNA (DNA complementare) oppure, conoscendo la sequenza amminoacidica della proteina codificata dal gene che ci interessa, è possibile assemblare direttamente un breve filamento di DNA sintetico.
Il processo di appaiamento della sonda al DNA di interesse è detto ibridazione.
In generale si forma DNA ibrido quando si appaiano filamenti di DNA (o di DNA ed RNA) di provenienza diversa.
Amplificazione DNA: PCR
La clonazione del DNA tramite proliferazione di un batterio nel quale è stato introdotto un vettore ricombinante (plasmide o fago) contenente il segmento genico di interesse non è l’unica modalità per ottenere un numero elevato di segmenti di DNA identici (amplificazione DNA). Il metodo più rapido ed economico per ottenere molte copie di una sequenza di DNA è infatti la reazione a catena della polimerasi (PCR). Si tratta di un sistema in vitro, che non prevede cioè l'uso di cellule ed è basato sull'attività dell’enzima DNA-polimerasi. Le DNA-polimerasi sono enzimi in grado di legare in successione ordinata i nucleotidi e di costruire quindi filamenti di DNA complementari ad un primo filamento che fa da stampo. Le DNA-polimerasi utilizzate attualmente sono tutte estratte da batteri che vivono in condizioni di temperatura estreme, in modo da essere termostabili. La "Taq-polimerasi" ad esempio, è in grado di lavorare ad alte temperature, fino anche a 92°C, temperatura alla quale avviene il processo di denaturazione del DNA. In passato questa DNA-polimerasi veniva estratta da Thermophilus aquaticus, un batterio che vive nelle acque termali calde. Attualmente viene invece prodotta in batteri geneticamente modificati per l'espressione e la sintesi dell'enzima.
La PCR prevede che il DNA venga riscaldato per dividere i due filamenti della doppia elica (denaturazione), premessa indispensabile per poterlo copiare. Se la polimerasi usata non fosse termoresistente, ad ogni ciclo si dovrebbe aggiungere nuovo enzima per sostituire quello inattivato dal calore. Il campione di DNA da amplificare viene posto in una soluzione contenente DNA polimerasi, nucleotidi (dNTP, desossinucleotidi trifosfati) e primer.
Un primer è un breve frammento di DNA a singola elica, un oligonucleotide di circa 20 basi, complementare alla porzione iniziale della sequenza di DNA che si vuole amplificare. Esso fornisce alla polimerasi un punto di innesco per la sintesi del nuovo filamento di DNA. Sono necessari due diversi primer, uno per ogni filamento, poiché dovranno posizionarsi all’estremità 3’ di ogni filamento (la polimerasi allunga il nuovo filamento solo in direzione 5’ → 3’). Il primer è aggiunto in grande eccesso al DNA affinché le due eliche parentali non si riappaino. La provetta con la miscela di reagenti è posta nella apparecchiatura per PCR, capace di cicli successivi di riscaldamento e raffreddamento a temperature specifiche per ogni reazione. Le fasi della PCR si possono così riassumere:
- denaturazione del DNA a temperature di 90-95°C
- attacco del primer (ibridazione) al sito bersaglio a temperature di 50-60°C
- fase di allungamento, a temperature di circa 70°C, ad opera della polimerasi, in cui l'enzima aggiunge nucleotidi all'estremità 3' del primer fino a completare la catena
Il ciclo si ripete in genere per 20-30 volte e termina con un raffreddamento a 4°C per bloccare ogni reazione.

Ad ogni ciclo il numero di molecole aumenta esponenzialmente. Dopo n cicli si formano 2n molecole di DNA.

La PCR risulta più rapida rispetto alla clonazione in vivo del DNA, ma a differenza di questa necessita della conoscenza delle sequenze nucleotidiche di ciascuna estremità del segmento di DNA che si vuole copiare.
Polimorfismo della lunghezza dei frammenti di restrizione (RFLP)
I siti di restrizione sono localizzati in punti differenti nel genoma di ogni individuo. Gli enzimi di restrizione tagliano perciò il DNA, producendo dei frammenti di restrizione che presentano lunghezze variabili da individuo ad individuo. Questa diversa distribuzione di frequenza della lunghezza dei frammenti in ciascun individuo è nota come polimorfismo della lunghezza dei frammenti di restrizione (RFLP Restriction Fragments Length Polymorphism). Si chiamano infatti polimorfismi le differenze ereditarie che si trovano tra gli individui in una popolazione.
Questa caratteristica può essere utilizzata per distinguere il DNA di un individuo da quello di qualsiasi altro. La distribuzione di frequenza delle lunghezze dei frammenti di restrizione è infatti caratteristica di ogni genoma.
Per separare i frammenti di diversa lunghezza e visualizzarne la distribuzione di frequenza si usano tecniche di elettroforesi.
L'elettroforesi è un metodo di separazione basato sulla diversa velocità di migrazione di particelle elettricamente cariche, depositate su di un opportuno supporto (carta o gel) se sottoposte ad un campo elettrico. L’elettroforesi è ad esempio impiegata nei comuni esami di laboratorio per quantificare le diverse classi di proteine del sangue.
Nel caso dei frammenti di restrizione, questi vengono depositati all’estremità di una lastra di gel. Poiché gli acidi nucleici hanno cariche negative, i frammenti migreranno verso il polo positivo. La velocità di migrazione dei frammenti di DNA è inversamente proporzionale al logaritmo della loro lunghezza. Al termine della corsa elettroforetica, DNA di lunghezza diversa sono risolti in bande di diverso peso molecolare (una sorta di codice a barre), che si possono visualizzare con sonde radioattive o fluorescenti.

In origine la tecnica RFLP fu messa a punto e viene ancora utilizzata per evidenziare un polimorfismo consistente nella presenza o assenza di un sito di restrizione, dovuto a mutazioni che vanno a colpire il sito di restrizione stesso facendolo scomparire se esiste, o creandone uno nuovo. Ad esempio, se l’allele di un difetto genetico genera rispetto a quello sano frammenti di restrizione diversi, è possibile usare questa tecnica per evidenziarne la presenza in un genoma.
Oggi però il termine ha acquisito un significato più ampio, poiché sono stati evidenziati dei tratti di DNA in cui, pur non essendovi mutazioni nei siti di restrizione, vi sono tuttavia polimorfismi di lunghezza dei frammenti.
Queste regioni altamente polimorfiche del genoma umano possono essere utilizzate per confrontare due campioni di DNA e verificare se provengono dal medesimo individuo (test legali e giudiziari in casi di delitti) o da individui imparentati (test di paternità)
L’impiego di enzimi di restrizione su DNA appartenenti a individui diversi determina infatti miscele di sequenze di DNA di lunghezze diverse e quindi tracciati elettroforetici diversi. Si parla di impronte digitali genetiche o DNA fingerprinting. L’analisi del DNA ricavato da sangue o altri tessuti consente di dirimere questioni legali, consentendo di attribuire ad una persona campioni biologici quali sangue, pelle, sperma, capelli e così via.
Nell'uomo, le regioni del DNA usate comunemente per ottenere il profilo genetico di un individuo si trovano su cromosomi 1, 2, 4, 5, 10 e 17 e sono costituite da regioni ripetitive o DNA in tandem o STR (Short Tandem Repeats) (microsatelliti e minisatelliti). Per sequenze ripetute in tandem si intende una stessa sequenza ripetuta molte volte di seguito, per esempio: ATGCC ATGCC ATGCC ATGCC , dove ATGCC è ripetuta in tandem quattro volte.
Si tratta di regioni altamente polimorfiche in lunghezza e amplificabili a partire da primer di circa 20 bp tramite PCR.
I microsatelliti consistono in sequenze ripetute in tandem in cui la sequenza ripetuta ha una lunghezza da 2 a 6 paia di basi, mentre nei minisatelliti l’unità ripetuta è composta da 10 a 200 paia di basi.
Dato che si conosce la sequenza amplificata e la natura della ripetizione, dalla lunghezza del frammento amplificato si può dedurre quante ripetizioni esso contenga, e quindi di quale variante si tratti.
Ciò che si confronta tra due campioni, sono le bande visibili su un gel e corrispondenti al frammento di restrizione marcato con una sonda radioattiva o luminescente.

DNA ricombinante e principio di precauzione
Molte persone, specialmente in Europa, considerano gli OGM un potenziale pericolo per l'ambiente e per la salute, sebbene esistano ormai migliaia di studi sulla loro sicurezza e non siano emersi dati significativi da un punto di vista scientifico.
Il primo OGM fu ottenuto nel 1973 da Stanley Cohen e Herbert Boyer (inserimento in E.coli del gene per la resistenza all’antibiotico canamicina prelevato da Staphylococcus aureus).
L’anno dopo la comunità scientifica si autoimpose una moratoria internazionale sull'uso della tecnica del DNA ricombinante. Questo permise di valutare lo stato della nuova tecnologia ed i possibili rischi, attraverso un approccio precauzionale. La conferenza che raccolse i risultati ottenuti si tenne ad Asilomar in California e concluse che gli esperimenti sul DNA ricombinante potessero procedere a patto che rispettassero severe linee guida, poi redatte dal National Institute of Health (NIH) ed accettate dalla comunità scientifica.
Queste linee guida sono tuttora la base che ispira tutte le ricerche di laboratorio che riguardano esperimenti di trasformazione genica e le normative che presiedono all'uso commerciale ed al rilascio ambientali di tali organismi nell'ottica di consentire l'utilizzo solo di varietà e animali che siano stati riconosciuti sicuri per l'ambiente e per il consumo umano e animale.
Ad oltre 30 anni dalla Conferenza di Asilomar la tecnica del DNA ricombinante ha mostrato più benefici che rischi. Ad oggi infatti non è noto alcun problema specifico legato ad essa.
Nonostante ciò in gran parte dell’opinione pubblica sono diffusi i timori che la manipolazione del genoma possa comportare delle conseguenze non previste, sia rispetto alla modificazione dell'alimento (e quindi alle conseguenze sulla salute umana) sia rispetto all'interazione con le altre specie viventi e quindi alle conseguenze sull'ambiente.
Contro la produzione e l’introduzione degli OGM viene normalmente invocato il cosiddetto principio di precauzione.
Il principio di precauzione trova la sua prima formulazione ufficiale nella “Dichiarazione di Rio”, durante la Conferenza sull'Ambiente e lo Sviluppo delle Nazioni Unite del 1992, che all'articolo 15 recita:
«Ove vi siano minacce di danno serio o irreversibile, l'assenza di certezze scientifiche non deve essere usata come ragione per impedire che si adottino efficaci misure di prevenzione della degradazione ambientale»
Successivamente l’Unione Europea, nel recepire queste indicazioni nella sua normativa, ne ha allargato il campo di applicazione alla politica di tutela dei consumatori, della salute umana, animale e vegetale, ribadendo che «Il ricorso al principio di precauzione trova applicazione qualora i dati scientifici siano insufficienti, inconcludenti o incerti e la valutazione scientifica indichi che possibili effetti possano essere inaccettabili e incoerenti con l'elevato livello di protezione prescelto dall'Unione europea»
La formulazione del principio di precauzione è tuttavia talmente ambigua e generica da consentire interpretazioni diverse ed opposte. Si tratta infatti di capire cosa debba intendersi per “assenza di certezze scientifiche” e per “efficaci misure di prevenzione”.
Nella sua applicazione più integralista al problema della tecnologia del DNA ricombinante, il principio di precauzione afferma che, poiché non esistono dati scientifici sufficienti ad affermare con certezza che gli OGM non sono dannosi per l’uomo e per l’ambiente, gli OGM non devono essere autorizzati. In altre parole si afferma che una mancanza di prove di dannosità non è sufficiente a garantire la non dannosità ("l'assenza di prove non è prova di assenza" disse Carl Sagan).
A ben guardare viene qui ribaltato uno dei cardini del diritto. La norma che recita “innocente fino a prova contraria” viene sostituita da “colpevole fino a prova contraria”.
Il problema è che la scienza, per sua natura, non è in grado di fornire risposte certe e questa “incertezza” risulta particolarmente pesante quando si tratta di descrivere e prevedere il comportamento di sistemi altamente complessi ed interagenti come sono appunto gli ecosistemi.
In questa situazione è ovviamente difficile capire quando si possa affermare di avere una quantità di dati sufficiente per esprimere un giudizio ponderato su una specifica tecnologia. In realtà, tutti i giorni si prendono decisioni, o si scelgono opzioni, in base a una ragionevole valutazione del rapporto rischi/benefici, dando un peso e un valore ai dati a seconda delle priorità definite dalle normative esistenti. Se si dovesse prendere una decisione solo sulla base della prova assoluta di assenza di rischio, qualsiasi società si bloccherebbe nel suo sviluppo tecnologico e sociale. Non esiste infatti azione umana che non comporti, in potenza, qualche rischio. L’applicazione del principio di precauzione risulta invece ragionevole quando viene applicato al fine di escludere la presenza, per le nuove tecnologie, di problemi potenzialmente maggiori rispetto a quelli esistenti per le tecnologie o prodotti che si intendono sostituire.
È inoltre da sottolineare che non esistono tecnologie, anche semplici, che non comportino possibili effetti collaterali, per cui il rischio zero non può essere usato come parametro di valutazione in quanto impossibile da accertare.
L’assenza di “certezze scientifiche” non significa comunque che, compatibilmente con lo stato attuale delle conoscenze, non sia possibile dare delle risposte a molti dubbi e timori che accompagnano l’introduzione degli OGM. È ovvio che se le nostre conoscenze in futuro si modificheranno, potranno di conseguenza modificarsi anche le nostre valutazioni su questa tecnologia.
Tra le varie fonti di preoccupazione le più frequenti riguardano la possibilità che gli OGM
- si comportino come specie invasive (superinfestanti), che si affermano nell'ecosistema a danno di altre specie e varietà,
- riducano la biodiversità.
- inducano resistenza agli antibiotici in microrganismi patogeni per l’uomo,
- modifichino il patrimonio genetico di chi se ne alimenta
- contaminino prodotti non-GM
- siano pratiche contro natura
- siano tossici o allergenici e quindi pericolosi per la salute
Vediamo le risposte che la scienza può dare
Le biotecnologie sono contro natura?
Dal dibattito sulle biotecnologie emerge spesso una condanna morale di questa tecnologia. La modifica diretta del DNA ha dato a molti l’impressione di violare un limite imposto dalla natura. Gli OGM sarebbero “contro natura”.
In realtà, la modifica del DNA degli organismi viventi a opera dell’uomo, ha origini antiche e coincide con la nascita stessa dell’agricoltura e della domesticazione di piante e animali. L’intervento umano è stato tanto rilevante da rendere la maggior parte delle piante coltivate e degli animali completamente inadatti a vivere negli ecosistemi naturali. Alcune di queste piante sono state modificate in modo così significativo da presentare una bassissima fertilità che ne limita la dispersione nell’ambiente (le banane coltivate vengono riprodotte esclusivamente tramite talea in quanto prive di semi). Alcune specie sono state incrociate con altre molto distanti geneticamente per trasferire geni utili. La segale è stata incrociata con il frumento per ottenere il triticale, un cereale che non esiste in natura. In questo contesto, se gli OGM sono da considerare innaturali, lo sono anche l’agricoltura e l’allevamento che nei secoli hanno invaso habitat prima dominati da specie selvatiche, rimpiazzandole con pochissime specie coltivate, frequentemente importate da altri continenti. Va aggiunto inoltre che le modifiche introdotte dall’uomo nel genoma di specie utili sono state per lo più casuali e inconsapevoli, con effetti sulla pianta, sull’ambiente e sull’uomo largamente non prevedibili e non ancora studiati, ma comunque considerati “normali”.
Le nuove tecnologie, mimando processi che avvengono già in natura (è possibile infatti ritrovare le tracce del trasferimento di DNA da batteri a piante e viceversa, così come in altri organismi), permettono modifiche mirate del DNA, entro certi limiti prevedibili e controllabili.
Gli alimenti GM possono modificare il DNA di chi se ne ciba?
Una persona ingerisce circa un grammo di DNA al giorno tramite l’alimentazione. A contatto con i tessuti dello stomaco il DNA viene ridotto in piccoli frammenti (in genere minori di 400 paia di basi). In alcuni esperimenti su ratto si è osservato come una piccola parte del DNA così frammentato (inferiore allo 0,1%) può essere assorbita dalle cellule dell’intestino e rimanere per qualche ora, prima di essere degradata, nel sangue o nel fegato, senza comportare conseguenze. In ogni caso il DNA transgenico non è chimicamente differente dal DNA non-transgenico. Se ne deduce che se potesse integrarsi al DNA umano, modificandolo, potrebbe farlo anche il DNA contenuto in qualsiasi altro alimento.
Gli OGM sono dannosi (tossici o allergenici) per la salute umana o animale?
Gli OGM, prima di ricevere l’autorizzazione alla coltivazione e alla commercializzazione, devono superare un elevato numero di test di sicurezza. I test richiesti partono dallo studio della nuova proteina prodotta dalla pianta transgenica. Il DNA inserito (transgene) si “esprime”, infatti, tramite la produzione di una proteina, responsabile della caratteristica desiderata (ad es. resistenza agli insetti, tolleranza a un erbicida, maggior contenuto di vitamina, ecc.).
Se i test danno esito negativo, cioè se la proteina non è tossica o allergenica, si passa a valutare la “sostanziale equivalenza” della pianta transgenica nei confronti di piante analoghe non transgeniche. La sostanziale equivalenza richiede test nutrizionali condotti su animali. Se da questa analisi non emergono differenze significative il nuovo OGM viene considerato “sostanzialmente equivalente” alle varietà convenzionali. Va aggiunto anche che i prodotti autorizzati vengono monitorati per almeno 3 anni negli Stati Uniti e per tutta la durata dell’autorizzazione nell’Unione Europea, al fine di riscontrare eventuali effetti indesiderati sulla salute o sull’ambiente.
Gli OGM possono diffondere resistenze agli antibiotici rendendo più difficile la cura delle malattie?
Alcuni OGM in commercio, oltre al gene di interesse, contengono un gene che conferisce la resistenza a un antibiotico. L’uso del marcatore di resistenza è necessario per verificare che il trasferimento genico sia avvenuto correttamente. Il 90% delle varietà transgeniche autorizzate hanno il gene che conferisce resistenza alla kanamicina. Il restante 10% è resistente ad altri due antibiotici: ampicillina e igromicina. Questi tre geni di resistenza sono comunque molto diffusi tra i microrganismi presenti naturalmente nei suoli. Inoltre, kanamicina e igromicina non sono utilizzati in medicina, a causa della loro tossicità. Per l’ampicillina la situazione è diversa: questo antibiotico è ancora utilizzato, sebbene il suo impiego sia in declino a causa della diffuse resistenze naturali (più del 50% degli enterobatteri è resistente a questo antibiotico). La probabilità quindi che un agente patogeno per l’uomo acquisisca il gene di resistenza dai batteri già presenti nell’intestino o nel suolo è di gran lunga superiore alla probabilità di acquisirlo da alimenti ricavati da piante transgeniche. Anche nel caso in cui questo trasferimento dovesse avvenire, non si tratterebbe comunque di resistenze ad antibiotici utilizzati comunemente in terapia. In ogni caso, la normativa europea ha stabilito che a partire dal 2006 non saranno più autorizzati OGM che contengano geni di resistenza agli antibiotici utilizzati in terapia.
Cos’è la “contaminazione” genetica?
È diventato uso comune parlare di “contaminazione” genetica quando vengono ritrovate tracce di OGM in sementi o prodotti non-GM. Il termine è fuorviante, perché suggerisce un’intrinseca pericolosità dei prodotti GM, non tenendo conto che gli OGM non possono essere commercializzati se non sono stati riconosciuti sicuri. Al fine di evitare fraintendimenti, sarebbe quindi più appropriato parlare di “presenza accidentale” di OGM in lotti di semi non OGM o in prodotti non OGM. Si è finora riscontrato che la presenza accidentale di OGM in prodotti non OGM è inferiore allo 0,1%, percentuale molto più bassa rispetto alle soglie normalmente tollerate in agricoltura per la presenza di varietà tossiche (come la colza HEAR – 2%). Pertanto, trattandosi di organismi di cui è stata verificata la non dannosità, non dovrebbero generare alcuna problematica aggiuntiva.
Le piante GM possono diventare infestanti?
Il presupposto per la comparsa di nuove piante infestanti è che una pianta acquisisca resistenza a insetti e malattie o che si modifichi fino a presentare una serie di nuovi caratteri di invasività (ovvero la capacità di diffondersi autonomamente anche fuori dal campo coltivato)
per mutazione spontanea o per ibridazione con altre piante selvatiche o coltivate.
Gli OGM, da quanto emerso dagli studi fino ad oggi pubblicati, non sono di per sé più infestanti delle colture convenzionali. Le piante coltivate infatti necessitano per sopravvivere di cure costanti (devono essere seminate, protette dalle malattie e dagli agenti avversi) e non disperdono il seme quando mature, come invece accade per le piante selvatiche ed infestanti.
È inoltre provato che quando vengono disperse nell’ambiente naturale tendono a scomparire in 2-3 generazioni (come dimostrato per mais, colza, patata e bietola. Gli ibridi ottenuti dall’incrocio tra una pianta selvatica (infestante o meno) e una pianta coltivata sono spesso poco vitali e possiedono una bassa fertilità. Nei casi in cui gli ibridi siano fertili e acquisiscano resistenze a malattie o ad altre avversità ambientali, essi potrebbero acquisire una maggiore invasività. Il rischio connesso è però il medesimo sia per le piante GM, sia per le piante non-GM. Un esempio spesso citato riguarda la resistenza agli erbicidi. L’eventuale ibridazione tra varietà resistenti (coltivate) e non resistenti (selvatiche o coltivate) potrebbe portare, infatti, alla creazione di piante infestanti resistenti all’erbicida, costringendo l’agricoltore a usare nuovi prodotti per il loro controllo.
Quale impatto hanno gli OGM sulla biodiversità?
L’impatto degli OGM sulla biodiversità è un tema complesso. Qualunque tipo di intervento umano o anche semplici eventi naturali hanno un impatto sulla biodiversità.
Da quando l’agricoltura esiste (circa 12-13.000 anni) l’uomo ha condotto scelte o selezioni tra i genotipi delle specie da lui ritenute interessanti. Il problema della riduzione della biodiversità non è quindi nuovo. La ricerca di piante ad alta produttività, per esempio, ha ridotto radicalmente il numero di specie utilizzate a scopo alimentare. Attualmente 20 specie vegetali forniscono il 90% del cibo umano e quattro di esse arrivano da sole al 50% (frumento, mais, riso e patata). Questo processo non viene necessariamente amplificato dagli OGM.
L’ecosistema agricolo è stato inoltre profondamente modificato dall’introduzione di coltivazioni esotiche. Colture originariamente tipiche dell’America, come mais, patata e pomodoro, o dell’Asia, come soia, riso e kiwi, sono diffuse anche nell’ambiente europeo. Inoltre, con l’introduzione di specie in territori diversi da quelli nativi è stata spesso introdotta anche la flora selvatica affine a tale specie, immettendo così nell’ambiente anche le potenziali infestanti (e spesso anche i relativi parassiti). La coltivazione della soia, ad esempio, richiede dei Rizobi, batteri del terreno necessari al suo sviluppo, ma poco ancora oggi si conosce sull’impatto, a livello di biodiversità del suolo, dello spargimento di tali batteri.
Chiedersi se gli OGM siano una minaccia per la biodiversità ha poco senso. Più corretto sarebbe infatti domandarsi se questi organismi influiscono sulla biodiversità in modo diverso rispetto a quanto accade per le colture convenzionali.
Ma valutare l’impatto sulla biodiversità dell’introduzione di una nuova varietà (GM o meno) in agricoltura è comunque estremamente difficile, trattandosi di sistemi altamente complessi.
Da una parte, ad esempio, vi è la possibilità che geni di varietà commerciali (GM o meno) possano entrare a far parte del DNA di specie sessualmente compatibili. Questo fenomeno naturale può in teoria portare ad una diminuzione di biodiversità se vengono trasferiti agli ecosistemi naturali geni in grado di aumentare la competitività di questi ibridi a scapito di specie selvatiche. Dall’altra parte dobbiamo considerare che la mancanza di scambio genico è per altri versi pericolosa per la biodiversità. In definitiva, non ci sono ragioni per ritenere che la diffusione di colture OGM debba essere considerata potenzialmente più dannosa, per la biodiversità, dell’introduzione di colture “tradizionali”.
Problemi etici
Naturalmente le tecniche del DNA ricombinante hanno sollevato anche numerosi problemi etici, come ad esempio i dubbi sulla liceità della manipolazione dei geni umani e della loro introduzione in altri organismi e/o dell’introduzione di geni estranei in cellule e organismi umani (es. per terapie geniche). Ovviamente su questi aspetti la scienza non ha e non può avere risposte.
Evoluzionismo
Il termine “evoluzionismo” è molto spesso utilizzato come sinonimo di “evoluzione biologica” e rimanda alla teoria darwiniana dell’evoluzione delle specie per selezione naturale. Qui in effetti tratteremo di tale teoria, tuttavia la teoria della evoluzione della vita rappresenta solo un caso particolare all’interno di un “approccio evoluzionista” alla visione ed all’interpretazione scientifica del mondo.
L'idea di una natura dinamica contrapposta a una natura statica è una visione molto antica che ha generato polemiche e controversie sin dal tempo della prima speculazione greca intorno al mondo. Le due scuole di pensiero si possono esemplificare da una parte attraverso Parmenide, che fonda la sua filosofia sull'essere, immutabile, e dall'altra con Eraclito per cui tutto scorre continuamente (pànta rèi).
Questi due opposti modi di interpretare la realtà hanno percorso la storia del pensiero occidentale fino ai nostri giorni. Credere che la natura sia in continua trasformazione significa spostare l'attenzione dallo studio del mondo a quello delle "leggi" che regolano il mondo. Gli unici aspetti invariabili nell’universo che cambia sono le leggi fisiche del cambiamento (attualismo). L'idea che l'universo, gli astri, la Terra, le montagne, i mari, le specie animali e vegetali col tempo si trasformino, nascano e muoiano oggi ci può sembrare scontata ma, storicamente, per affermarsi ha dovuto sconfiggere un paradigma (= un modello esplicativo) fissista e creazionista molto forte, secondo il quale l'universo è stato fatto da Dio in modo definitivo e statico ed ogni cambiamento è un evento eccezionale e spesso catastrofico (teoria del catastrofismo) all’interno di un mondo fondamentalmente stabile.
Il concetto di evoluzione, però, contiene molto di più della semplice constatazione che tutto cambia e si trasforma, contiene anche una spiegazione di come avviene il cambiamento. L’affermarsi di concezioni evoluzionistiche è inoltre strettamente legata alla “scoperta del tempo profondo”. Per gli uomini del Seicento la terra non aveva più di 6000 anni. Nel 1650 James Ussher, arcivescovo anglicano di Armagh, integrando considerazioni storiche, conoscenze sui cicli astronomici e diverse fonti di cronologia biblica, calcolò che la Genesi era iniziata la sera del 22 Ottobre 4004 a.C. e terminata il giorno dopo, Domenica 23 Ottobre (Adamo ed Eva erano stati cacciati dal Paradiso Terrestre lunedì 10 Novembre 4004 a.C., il Diluvio Universale avvenne nell’anno 2349 a.C e l’arca di Noè approdò sul monte Ararat il Mercoledì 5 Maggio 2348 a.C).
La difficoltà di concepire la storia della Terra secondo intervalli di tempo più estesi rendeva ovviamente difficoltosa una spiegazione di tipo evolutivo dei fenomeni naturali. L’evoluzionismo interpreta infatti i fenomeni del presente come il risultato del succedersi di piccole e lentissime variazioni, talmente lente da essere impercettibili per il singolo osservatore, ma che in tempi lunghissimi possono generare enormi cambiamenti.
Montagne, fiumi, stelle, pianeti, animali e piante hanno sempre avuto il medesimo aspetto, a memoria d’uomo e se la Terra ha solo 6000 anni è abbastanza comprensibile che il ritrovamento di fossili appartenenti ad organismi estinti e le grandi stratificazioni rocciose che sembravano indicare condizioni fisiche, chimiche e biologiche passate, completamente diverse da quelle attuali, venissero interpretate come la conseguenza di enormi e repentine catastrofi naturali.
Pensare la storia naturale come un lento processo evolutivo comportava dunque necessariamente una dilatazione del tempo.
Particolarmente interessante, da questo punto di vista, è il calcolo sull’età della Terra effettuato dal naturalista francese George-Louis Leclerc, Conte di Buffon (1707-1788). Nel 1778 Buffon ipotizzò che il sistema solare e quindi la Terra stessa si fossero formati a seguito di una violenta collisione tra una cometa ed il Sole, causando l'emissione di una serie di "globi" di fuoco che raffreddandosi avrebbero dato origine ai futuri pianeti e satelliti. Dalla fonderia di sua proprietà, fece costruire dieci sfere di ferro di diametro variabile e sulla base del tempo necessario affinché queste sfere si raffreddassero, dopo essere state riscaldate, ed ipotizzando l'interno della Terra costituito nella quasi totalità da ferro, egli fece una stima dell'età della Terra intorno ai 75.000 anni.
Nella seconda metà del ‘700 l’origine organica dei fossili era ormai comunemente accettata e le scoperte paleontologiche avevano ormai provato che esseri vissuti in passato erano scomparsi. L’immagine di un mondo immutabile e relativamente giovane era stata ormai definitivamente compromessa. Le collezioni naturalistiche di principi e signori si arricchivano di fossili appartenenti ad animali “preadamitici”.
L'interpretazione dei fossili fu d'altra parte un problema più volte affrontato nel passato Già alcuni pensatori greci avevano correttamente compreso che i fossili rappresentavano resti di organismi viventi poi pietrificatisi. E per quanto Leonardo Da Vinci, in alcuni scritti rimasti per altro sconosciuti, ed altri studiosi dopo di lui (Girolamo Fracastoro – 1517, Gerolamo Cardano - 1552, Bernard Palissy - 1580, Robert Hooke - 1665, N. Steensen (Stenone) - 1669) ne avessero chiaramente sostenuto l'origine organica, i più ritenevano trattarsi di concrezioni inorganiche (lapides figurati), rocce che la natura aveva foggiato a forma di pesce o di conchiglia in virtù di una "forza plastica" (aura seminalis, aurea petrifica, spiritus lapidificus, succus lapidescens) legata agli influssi astrali, seguendo in questo un errore aristotelico che pesò sul pensiero scientifico fino alla prima metà del settecento.
In alternativa furono date le spiegazioni più fantasiose, da quella secondo cui le "pietre figurate" non erano altro che resti di pasti degli antichi, alla teoria della "vegetazione delle pietre", secondo cui queste germogliano come le piante, fino ad invocare scherzi di natura (lusus naturae) o, come molti credevano, semplicemente i resti del diluvio universale.
Il 1785 fu una tappa decisiva nella storia del pensiero evoluzionistico e nella conquista del “tempo profondo” grazie all’opera del geologo scozzese James Hutton.
James Hutton e l’uniformismo
James Hutton (1726-1797) è ritenuto il fondatore della moderna geologia. A lui si devono intuizioni e scoperte fortemente innovative per i tempi che egli presentò nel suo Theory of the Earth (1785). I punti di maggior interesse della sua teoria sono
- La storia della Terra può essere spiegata attracerso una teoria unitaria sulla base del ripetersi di semplici fenomeni geologi che si avvicendano uniformemente e ciclicamente in tutti i momenti della storia geologica e in tutte le parti del globo (uniformismo), escludendo il ricorso ad eventi straordinari o miracolosi. Il sistema di Hutton tendeva a immaginare la Terra come una newtoniana “macchina perfetta”, regolata da leggi proprie ed immutabili, una macchina autoregolantesi e funzionante in tempi senza principio e senza fine, come l’universo degli astronomi
- Questi processi richiedono tempi lunghissimi. Il tempo geologico deve essere di gran lunga superiore alla capacità di immaginazione umana, non essendo possibile identificare le tracce né dell’inizio né della fine dei processi geologici.
- Fonda la teoria plutonista in contrapposizione alle ipotesi nettunistiche al tempo prevalenti.
La teoria nettunista di Werner assegnava un’origine marina a tutte le rocce. Solo più tardi Werner aggiunse alla sua classificazione le “serie vulcaniche”, comprendenti però solo alcune colate di lava, ritenute il prodotto recente, e poco importante, della “cottura” di strati carboniosi (“fuochi superficiali per autocombustione dei carboni”). La teoria postulava un grande Oceano primitivo circondante una terra emersa di piccole dimensioni. Il ritiro progressivo di questo mare avrebbe determinato l’emergere di nuove terre e la conseguente deposizione di tutti i materiali della crosta terrestre.
Le ricerche e le osservazioni eseguite da Hutton in Scozia gli avevano reso evidente che non solo il granito era una roccia ignea (magma fuso solidificatosi), ma che era stato in grado di penetrare gli strati sedimentari sovrastanti (intrusioni). Osservando strati sedimentari verticali sovrastati da strati orizzontali (discordanza stratigrafica) Hutton rimase impressionato da ciò che quella immagine gli suggeriva: il susseguirsi di mondi scomparsi, con gli strati verticali prodotti dalla sedimentazione di un antico mare, sollevati ed inclinati, poi erosi, poi di nuovo sommersi e ricoperti da altri strati sedimentari; a loro volta sollevati ed erosi. Hutton vedeva la terra come un pianeta dinamico che funzionava come una macchina azionata dal calore endogeno (da qui il termine plutonismo) e intuì che nessuna sorgente di calore superficiale sarebbe stata in grado di produrre i fenomeni vulcanici. In un ciclico ripetersi i fiumi erodevano i continenti e trasportavano i sedimenti negli oceani, mentre il calore profondo espandeva il pianeta, sollevando di nuovo i depositi marini compattati e li ritrasformava in continenti. Il sollevamento dei continenti (orogenesi) avrebbe prodotto strati inclinati e distorti. Una notevole anticipazione della moderna geodinamica e di ciò che noi oggi chiamiamo ciclo delle rocce.
Per i nettunisti la Terra non è molto antica. Le rocce, secondo Werner, sono state deposte, come precipitati chimici e come detriti sedimentari, come conseguenza del ritiro graduale di un Oceano primordiale (regressione), in un ordine ben preciso: a cominciare dai graniti e dagli gneiss primitivi fino ai più recenti strati sedimentari detritici. Secondo Hutton ed i plutonisti la Terra potrebbe invece aver attraversato un numero indeterminato di cicli di sollevamento e di erosione.
Hutton insiste su trasformazioni lente e regolari che hanno agito in passato ma sono tuttora operanti. continenti attuali sono soltanto formazioni transitorie nell'immensità del tempo geologico. Non esistono vestigia di un inizio né prospettive di una fine.
In questo senso va interpretata la sua affermazione “il presente è la chiave per spiegare il passato” ed il suo atteggiamento non va confuso con l’“attualismo” che verrà più tardi introdotto da Lyell. La filosofia di Hutton è piuttosto quella dell’uniformismo ed in questo senso il suo modo di pensare (come quello di Newton, del resto) è fortemente venato di teismo. Come non rischiare di confondere, infatti, un Universo perfetto, eterno e autosufficiente con la figura stessa della Divinità?
Non a caso, anche per questo, e non solo per le sue affermazioni sulla antichità della Terra, Hutton fu aspramente accusato di eresia.
Charles Lyell e l’attualismo
Charles Lyell (1797-1875) raccolse ed approfondì la lezione uniformista di Hutton, argomentandola in modo dettagliato nei Principles of Geology (3 vol 1830-1833) che ebbero una influenza determinante sullo stesso Darwin, tanto che questi ne testimoniò più volte nei suoi scritti. Lyell anticipò, tra l’altro, pur non traendone le debite conseguenze, quasi tutti i principi cui Darwin dette forma nell’evoluzionismo, a cominciare da quella “guerra di natura” che rimanda alla darwiniana “selezione naturale”. In geologia introdusse il principio dell’attualismo (gli eventi del passato si spiegano con forze ancora operanti e osservabili), principio che ricorda l’huttoniano “il presente è la chiave per spiegare il passato”. Ma se per Hutton questo implicava “uniformità dei fenomeni” (di oggi e del passato), per Lyell è piuttosto uniformità delle leggi di natura.
Secondo Lyell le modificazioni avvenute sulla Terra nelle ere geologiche passate dovevano essersi verificate con le stesse modalità e con la stessa intensità degli eventi geologici osservabili nel presente. Il passato geologico può dunque essere compreso meglio in termini di processi naturali che ancora oggi possiamo osservare, quali sedimentazione nei corsi d’acqua, erosione eolica ed idrica, avanzamento o ritiro dei ghiacciai, vulcanesimo e sismicità, Non è necessario invocare catastrofi né ipotizzare che i processi attuali abbiano agito con maggiore intensità in tempi precedenti. I cambiamenti sono lenti e costanti (gradualismo) e se gli effetti sono stati assai pronunciati ciò è dovuto all’accumularsi, in tempi estremamente lunghi, di tanti piccoli eventi. Il fattore tempo è di fondamentale importanza poiché anche piccole variazioni, sommate per intervalli di tempo sufficientemente lunghi, possono produrre effetti enormi.
Il gradualismo era in palese contrasto con la teoria del catastrofismo, avanzata nel 1815 dal naturalista francese Georges Cuvier (1769-1832). Cuvier fu uno dei più celebri studiosi di fossili dell’epoca, fondatore della paleontologia dei vertebrati e giustamente famoso per le sue ricostruzioni di animali preistorici a partire da pochi frammenti ossei. Riteneva che i fossili appartenenti a specie non più esistenti, testimoniassero antiche estinzioni avvenute come conseguenza di catastrofi, la più recente delle quali era appunto il Diluvio.
Già alla fine del ‘700 William Smith, un ingegnere minerario, aveva scoperto che strati rocciosi diversi potevano esser distinti in base al loro contenuto fossilifero, anche nel caso in cui fossero simili per litologia. Cuvier interpretò queste brusche discontinuità tra i fossili di uno strato ed i fossili del successivo come una prova dell’esistenza di eventi catastrofici.
Cuvier combattè aspramente sia il gradualismo di Lyell che, ovviamente, le nascenti teorie dell’evoluzionismo biologico (lamarckismo). I punti principali delle sue critiche sono i seguenti:
- Gli organismi raffigurati negli affreschi egiziani sono gli stessi presenti attualmente, come testimoniano i reperti mummificati ritrovati nelle tombe egizie.
- Le varietà e le razze che si osservano in natura non originano specie nuove e quelle prodotte attraverso l'allevamento, se tornate allo stato selvatico, tendono a riassumere la forma originaria
Evoluzione biologica: i precursori
Verso la fine del ‘700 la maggior parte degli scienziati accettava ormai l’idea che la complessa storia della vita sulla terra si fosse necessariamente svolta nell’arco di tempi lunghissimi e che i fossili di organismi di cui non si aveva il corrispondente attuale, rappresentassero le vestigia di specie estinte. L’idea di una evoluzione biologica degli esseri viventi cominciava dunque ad avere un terreno fertile su cui crescere. Tuttavia, come abbiamo già avuto modo di dire, c’è differenza tra ipotizzare un evoluzione biologica e fondare una teoria evolutiva che contenga al suo interno un meccanismo esplicativo delle modalità attraverso le quali l’evoluzione è avvenuta.
In questo senso le uniche due teorie evolutive avanzate furono il lamarckismo e il darwinismo. Ciò non toglie che l’idea di un’evoluzione biologica fosse precedente, anche se l’idea che risultò dominante fino alla prima metà dell’Ottocento fu quella della fissità delle specie, cioè della loro immutabilità e della loro formazione per intervento divino (creazionismo fissista). Il fissismo fu codificato dal naturalista svedese Carlo Linneo che nel 1751, nella sua opera Philosophia botanica, affermava che le forme viventi furono create per progetto divino e disposte in un ordine che si adattava a essere classificato (“Species tot sunt diversae quot diversas formas ab initio creavit infinitum Ens”).
Il naturalista svedese ebbe il merito di creare un valido sistema di classificazione binomia (nome generico e nome specifico, Homo sapiens, Drosophila melanogaster) dei viventi. Le sue idee creazioniste determinarono tuttavia una resistenza della comunità scientifica alle ipotesi evoluzioniste.
In un epoca in cui le verità scientifiche non potevano contraddire quelle religiose, il creazionismo fissista si accompagnava ad una visione finalistica della natura. Il finalismo della natura, il fatto cioè che gli esseri viventi possiedano delle strutture perfettamente adeguate per il compimento di certi scopi (gli uccelli hanno le ali per poter volare, i pesci le branchie per poter vivere nell'acqua e così via) era spiegato con l'intervento di Dio che aveva progettato la natura in modo perfetto (viviamo nel migliore dei mondi possibili). L'anatomia comparata, che studia le omologie tra le differenti specie - per esempio le ali degli uccelli, gli arti dei mammiferi e le pinne dei pesci - metteva in luce come Dio avesse utilizzato un solo progetto che aveva variato in tutti i modi possibili.
Scienziati come P. L. M. de Maupertuis (1698-1759) e filosofi come D. Diderot (1713-1784) erano tuttavia giunti ad una spiegazione che rovesciava l'apparente finalismo della natura. Le strutture degli animali sono perfette perché gli animali imperfetti e i "mostri" non sono sopravvissuti. La natura non è perfetta, i "mostri" nascono tutti i giorni, ma non possono sopravvivere né riprodursi (notevole anticipazione della “selezione naturale” di Darwin). Sviluppando queste considerazioni Maupertuis e Diderot arrivarono persino a prendere in considerazione l'idea di un prototipo di tutti gli animali che si sarebbe con il tempo variato in tutti i modi originando tutte le forme di vita presenti nel mondo.
Negli stessi anni, G. L. L. de Buffon (1707-1788), nella sua imponente Storia Naturale in 36 volumi, descriveva gli esseri viventi sottolineandone la trasformabilità. In polemica con Linneo, credeva che la classificazione degli esseri viventi fosse soltanto una comoda convenzione dell'uomo che non rispecchiava la realtà della natura. Le specie, infatti, avevano una storia di incroci, di parentele, di estinzioni, di cambiamenti avvenuti a seconda del clima e dell'ambiente. Non era quindi possibile classificarle come qualcosa di immutabile e fisso. Se Buffon non accettò mai l'ipotesi di un progenitore comune per tutti gli animali, parlava però di un piccolo numero di specie originarie da cui tutte le altre sarebbero derivate.
Erasmus Darwin (1731-1802), nonno del più celebre Charles, in Zoonomia, la sua opera principale, formulò delle ipotesi sulla trasformazione del vivente analoghe a quelle successivamente formulate da Lamarck. In molti casi si possono leggere affermazioni che anticipano il pensiero del nipote: "... sembra che lo scopo ultimo della lotta fra maschi sia che l'animale più forte e più attivo possa riprodursi...".
Lamarck e l’ereditarietà dei caratteri acquisiti
Ma la prima teoria articolata e coerente dell’evoluzione biologica si deve a J. B. P. A. Lamarck (1744-1829). Le sue ipotesi furono pubblicate per la prima volta nella Philosophie zoologique (1809) e poi riesposte in molti scritti successivi.
Anche Lamarck, come Cuvier ed altri, aveva osservato come gli strati di roccia più profondi e quindi verosimilmente più antichi racchiudevano fossili di forme viventi più semplici, ma, a differenza di Cuvier, ritenne che ciò potesse essere spiegato ipotizzando una sorta di derivazione degli organismi più complessi e più recenti da quelli più semplici e più antichi.
Questa derivazione biologica o evoluzione, come viene oggi chiamata, dipendeva per Lamarck da due fattori:
- i nuovi caratteri compaiono negli esseri viventi in risposta alla necessità ed ai nuovi bisogni indotti da un ambiente in continua trasformazione. I nuovi caratteri si sviluppano con l'uso, mentre regrediscono e scompaiono se inutilizzati (principio dell’uso e non uso). Poiché era opinione corrente che i vegetali non avvertissero sensazioni di bisogno, Lamarck non applicò la sua teoria al regno vegetale, indebolendola notevolmente.
- i nuovi caratteri acquisiti vengono trasmessi per eredità alle generazioni successive (principio dell’ereditarietà dei caratteri acquisiti).
Il suo esempio più famoso è quello della giraffa. Secondo Lamarck la giraffa moderna si è evoluta da antenati a collo corto, costretti ad allungarlo per raggiungere le foglie più alte dei rami in periodi di scarsa disponibilità alimentare. Questi antenati trasmisero il collo allungato ai loro discendenti i quali continuarono ad allungarlo e così via.
Nell'opera di Lamarck sopravvive la convinzione di origine aristotelica che l'uomo si trovi all'apice di una scala naturale. In questo senso l'evoluzione di una specie in un'altra più progredita sarebbe in qualche modo collegata ad un principio creativo finalizzato alla comparsa dell'uomo sulla terra.
Una specie di principio vitale universale che, sotto forma di una forza interna agli esseri viventi, tende a trasformarli in forme organiche più evolute e sempre più complesse, trovando la propria realizzazione nella specie umana. La teoria lamarckiana è in questo senso ancora fortemente finalistica.
Per giustificare il fatto che, nonostante il processo evolutivo trasformi gli organismi più semplici in organismi più complessi, i gradini più bassi della scala naturale continuano ad essere occupati da forme di vita estremamente semplici, Lamarck accetta l’ipotesi aristotelica della generazione spontanea degli organismi più semplici dalla materia organica inanimata.
Alla luce delle attuali conoscenze di genetica possiamo affermare che un essere vivente può effettivamente perfezionare con l'uso un certo organo come, d'altra parte, può giungere all'atrofia di certe funzioni se non adeguatamente utilizzate, ma ogni modificazione dell'organismo che non si rifletta in una corrispondente modificazione del corredo cromosomico non può essere ereditata e non può di conseguenza dar luogo a nessuna forma di evoluzione.
I caratteri acquisiti sono modificazioni del fenotipo e non del genotipo.
La teoria lamarckiana non fu accolta favorevolmente dalla critica tradizionalista, trovando un convinto oppositore proprio in Cuvier che, col suo crescente prestigio nel campo degli studi biologici e paleontologici, ne decreterà il declino. Morti Lamarck e Cuvier, si spensero gli echi dello scalpore suscitato dalla teoria lamarckiana. Bisognerà aspettare un trentennio perché, con Darwin (1809-1882), attraverso una più completa e coerente documentazione, possa essere conferito al concetto di evoluzione biologica la dignità di teoria scientifica e di paradigma fondamentale della biologia moderna, avviando così alla crisi tutta una tradizione di pensiero.
Darwin e la selezione naturale
Nonostante la teoria lamarckiana non sia oggi più sostenibile, essa rappresentò il primo tentativo coerente di superare il concetto di fissità delle specie in favore di una visione evoluzionistica del mondo dei viventi e rappresentò un punto di riferimento fondamentale per l'opera di Darwin.
Charles Darwin e Alfred Russel Wallace formularono contemporaneamente e separatamente una teoria evolutiva degli esseri viventi per selezione naturale.
Nel 1859 Darwin pubblicò, dopo un lavoro di elaborazione durato circa venti anni "On the origin of species by mean of natural selection", un'opera fondamentale che rivoluzionò ogni precedente teoria.
Darwin conosceva le teorie di Lamarck. L'originalità del suo lavoro non sta quindi nell'aver proposto una teoria evolutiva, ma nell'aver dato un'interpretazione scientificamente soddisfacente dei meccanismi evolutivi.
Oltre alle ipotesi di Lamarck, fu fondamentale per la formulazione della teoria darwiniana la conoscenza del lavoro dell'economista inglese Thomas R. Malthus. "Saggio sul principio di popolazione" (1798) e delle ipotesi attualiste sostenute dal geologo inglese Charles Lyell che, nei suoi "Principi di geologia", ripropose con maggior concretezza scientifica la teoria huttoniana portando a suo favore nuove prove ed argomentazioni.
Nel suo "Saggio" Malthus afferma che mentre la popolazione umana si sviluppa secondo una progressione geometrica quindi con andamento esponenziale (y = ax), le risorse ed i beni di consumo crescono seguendo una progressione aritmetica (y = bx). E' quindi inevitabile, secondo Malthus, che si arrivi ad una situazione in cui le risorse non sono sufficienti per tutta la popolazione.

Malthus utilizza tale analisi per giustificare "scientificamente" l'esistenza dei poveri e per dimostrare "l’inevitabilità della miseria" in un paese in cui la rivoluzione industriale ed un capitalismo non ancora temperato da uno stato sociale avevano creato enormi ingiustizie sociali.
AI di là delle implicazioni sociali ed economiche dell'analisi malthusiana, interessa qui, sottolineare come l'approccio di Malthus sia riconducibile ad una concezione biologica della società, vista come un'entità soggetta alle stesse leggi naturali che regolano la riproduzione e la moltiplicazione di una qualsiasi popolazione di organismi vegetali o animali.
Darwin intuisce che le conclusioni di Malthus, secondo cui il cibo e le risorse sono un fattore limitante per la crescita delle popolazioni, erano vere per tutte le specie e non solo per la specie umana.
Ad esempio Darwin calcolò che una coppia di elefanti, che sono animali con uno dei più bassi tassi riproduttivi, sarebbe in grado di generare in 750 anni circa 19 milioni di individui. Poiché invece il numero degli elefanti rimane mediamente lo stesso da qualche centinaio d'anni a questa parte, Darwin ne dedusse che gli elefanti attualmente presenti devono essere stati in qualche modo "scelti" tra tutti quelli che si sarebbero potuti generare.
Il processo attraverso il quale gli esseri viventi sono "scelti" per perpetuare la loro specie è detto da Darwin selezione naturale.
Ma lo studioso che più di ogni altro influenzò Darwin fu probabilmente Charles Lyell. Lyell infatti portò nuovi numerosi argomenti a favore della teoria dell'attualismo. In realtà Lyell rimase antievoluzionista fino al 1862. Tuttavia, ciò di cui la teoria di Darwin aveva bisogno erano tempi geologici sufficientemente estesi e questo era proprio quello che la teoria di Lyell gli offriva.
Se la terra, come affermava Lyell, possiede una lunga storia fatta, non di catastrofi, ma di impercettibili variazioni che si sommano e si accumulano trasformando lentamente l'ambiente geologico, è naturale e giustificato ritenere che anche il mondo biologico abbia alle sue spalle una lunga storia di graduali modificazioni.
Tuttavia l'esperienza probabilmente decisiva Darwin la fece durante un lungo viaggio di circumnavigazione della terra sul Beagle, dal quale trasse un'enorme quantità di osservazioni sulla lussureggiante e multiforme flora e fauna tropicale.
Dopo un lavoro di elaborazione durato più di vent’anni, Darwin sintetizzò le sue osservazioni in una teoria organica dell'evoluzione, basata sulla selezione naturale, i cui punti essenziali possono essere così riassunti:
- tutti gli organismi producono una progenie di gran lunga superiore alle capacità che l'ambiente ha di sostenerla. Per questo motivo tra tutti gli individui che nascono solo alcuni sono in grado di raggiungere la maturità ed hanno la possibilità di riprodursi;
- in ogni generazione sono presenti individui casualmente molto diversi gli uni dagli altri, con una grande variabilità di caratteri;
- non tutti i caratteri presenti in ciascuna generazione hanno la stessa importanza rispetto alla possibilità di sopravvivenza dell'individuo relativamente all'ambiente in cui vive. Alcuni caratteri possono rendere l'individuo che li porta più adatto a sopravvivere in quel determinato ambiente. Altri caratteri possono risultare indifferenti o addirittura dannosi o letali;
- tra gli individui di una generazione solo quelli che presentano casualmente i caratteri che li rendono più adatti sopravvivono ed hanno così la possibilità di prolificare. Ciò consente loro di trasmettere i loro caratteri alla discendenza, mentre i caratteri degli individui inadatti si estinguono con essi.
L'ambiente si comporta cioè, secondo Darwin, come un filtro in grado di selezionare, di generazione in generazione, solo gli organismi viventi più adatti alla sopravvivenza.
Darwin capovolge in questo modo la teoria di Lamarck. Non è l'ambiente che induce dei cambiamenti evolutivi negli organismi, ma tali cambiamenti sono già presenti casualmente in ogni generazione sotto forma di variabilità individuale. L'ambiente attua solo una selezione su tale variabilità.
Per Darwin l’ambiente gioca un ruolo puramente passivo di filtro, mentre per Lamarck esercita un ruolo attivo, inducendo la variazione dei caratteri.
Poiché non tutti gli individui possono giungere a maturità essendo le risorse naturali insufficienti, si instaurerà una competizione tra gli individui per la conquista delle risorse, la lotta per la sopravvivenza mutuata dalla lettura di Malthus.
A questo punto saranno "naturalmente selezionati" e sopravviveranno, perpetuando così i loro caratteri, solo gli individui più adatti a quel determinato ambiente.
Per Darwin, che aveva una certa conoscenza dei sistemi di incrocio adottati dagli allevatori, la selezione naturale era del tutto analoga al processo di selezione praticato per il bestiame. Nel caso della selezione artificiale gli uomini scelgono gli esemplari di piante o di animali da far riprodurre in base alle caratteristiche che sembrano loro più utili; nel caso della selezione naturale l'ambiente si sostituisce all'uomo.
Inoltre, secondo Darwin, le variazioni tra individui, che sono presenti in ogni popolazione naturale, sono dovute unicamente al caso. Per giustificarle non è possibile chiamare in causa ne' l'ambiente, ne' una forza vitale, ne' un qualche inconscio impulso dell'organismo.
In sè le variazioni non hanno ne' scopo ne' direzione. È l'azione della selezione naturale su parecchie generazioni che dà, in modo del tutto casuale, la direzione all'evoluzione.
Molto spesso si sente affermare che la grande rivoluzione del darwinismo è connessa all'ipotesi di un'evoluzione dell'uomo da esseri inferiori.
In realtà l'evoluzionismo era una teoria di cui si parlava ormai da quasi un secolo e molto probabilmente il forte impatto della teoria darwiniana non è da ricercarsi nella negazione del creazionismo biblico, ma nella dimostrazione dell’inutilità di un disegno divino e più generale di un progetto in natura.
Lo sviluppo della civiltà occidentale è pervaso dall'idea che il mondo naturale e l'uomo in particolare siano l'espressione di un progetto intelligente (si pensi al Demiurgo del Timeo platonico). All'idea di un progetto in natura si può iscrivere anche la convinzione aristotelica dell'esistenza per ogni specie di un tipo perfetto, cui i vari membri della specie si avvicinavano come copie imperfette.
Gli stessi evoluzionisti predarwiniani, come Buffon, E. Darwin e Lamarck, non abbandonarono mai l'idea di un progetto sotteso al processo evolutivo. L'evoluzione per essi non era governata dal caso, ma dalla tendenza a raggiungere la perfezione.
L'evoluzione biologica è in sè compatibile con l'idea di progetto. Si può ad esempio sempre ammettere che Dio abbia creato l'uomo gradualmente, servendosi di un meccanismo evolutivo da lui messo in moto e controllato. La selezione naturale proposta da Darwin è invece un meccanismo assolutamente casuale e adirezionale, che esclude la necessità di un progetto in natura.
L'avversione della Chiesa per la teoria darwiniana fu immediata e per alcuni versi ancora più forte di quella espressa in precedenza nei confronti del copernicanesimo. Nel 1543 l'astronomo polacco aveva dato il colpo di grazia al dogma geocentrico spostando la Terra in zona periferica, ora Darwin lo assestava a quello antropocentrico, relegando l’uomo nel regno animale. Così se in una delle sue rare visite al British Museum lo scienziato venne indicato da un pastore anglicano presente in quel luogo come “l’uomo più pericoloso d’Inghilterra”, anche molti scienziati suoi amici fra cui il geologo Lyell (le cui idee avevano ispirato il giovane Darwin) non accettarono la nuova teoria. Lyell dichiarò candidamente che preferiva considerare l’uomo come un arcangelo decaduto piuttosto che un essere derivato dalla scimmia, anche se in verità Darwin non disse (né avrebbe potuto dirlo) che l’uomo discende dalla scimmia. Nell’Origine delle specie l’autore non fa mai riferimento diretto al posto dell’uomo nell’evoluzione ma ai benpensanti non sfuggì che anche l’uomo, come tutti gli animali più evoluti, sarebbe dovuto derivare da una forma inferiore di vita che nel suo caso non poteva che essere la scimmia. Solo dodici anni più tardi Darwin scrisse l’Origine dell’uomo in cui avrebbe affrontato l’argomento specifico.
Darwin fu attaccato nel punto più debole della sua teoria, quello dell'eredità genetica. Egli non era infatti in grado di spiegare la natura della variabilità individuale ed il modo in cui essa veniva trasmessa di generazione in generazione.
Darwin riteneva, come la maggior parte dei biologi ottocenteschi, che i caratteri dei genitori venissero trasmessi ai figli, mescolandosi (eredità per mescolamento). Tale modello ha tuttavia delle implicazioni fatali per una teoria evolutiva che dipenda dalla variabilità. Infatti il mescolamento dei caratteri tende irrimediabilmente a generare individui intermedi, creando uniformità all'interno delle popolazioni nell'arco di poche generazioni e togliendo quindi la necessaria variabilità su cui agisce la selezione naturale. Darwin fu dunque costretto ad ipotizzare che la variabilità si rigenerasse grazie alla possibilità degli individui di trasmettere ai figli le nuove caratteristiche che acquisivano durante la loro esistenza.
Nel 1868, in "The variation of animals and plants", avanzò pertanto una “ipotesi provvisoria di pangenesi, teoria peraltro già presente nel Corpus hippocraticum, secondo la quale da tutte le parti dell'organismo si staccherebbe minuscole "gemmule", le quali andrebbero a confluire negli organi riproduttivi. La teoria, era strutturata in modo tale che le influenze ambientali che agivano sull'intero organismo modificandolo potessero modificare anche le gemmule riproduttive andando a far parte del patrimonio ereditabile.
In tal modo anche Darwin, paradossalmente, fu costretto ad ammettere, come Lamarck, l'ereditabilità dei caratteri acquisiti.
D'altra parte il lavoro pionieristico di Mendel sull'ereditarietà dei caratteri non ebbe sufficiente diffusione e Darwin non ne venne mai a conoscenza.
Verso la fine dell’Ottocento l'ereditarietà dei caratteri acquisiti si rivelò falsa, quando nel decennio 1880-1890 August Weismann tagliò la coda a topi che poi faceva accoppiare. Dopo venti generazioni i topi nascevano ancora con la coda, lunga quanto quella dei topi della prima generazione (in effetti il naturalista tedesco avrebbe potuto risparmiarsi una tale dimostrazione: sarebbe stato sufficiente osservare che le donne, nascono sempre con l’imene intatto a dispetto del fatto che, da Eva in poi, venga perso ad ogni generazione).
L'evoluzionismo post-darwiniano
Su tale argomento si iniziò a vedere chiaro solo all'inizio del '900, con la ripresa degli studi di genetica, quando Hugo de Vries formulò la sua teoria delle mutazioni, dove sostenne che nuovi caratteri possono improvvisamente comparire per modificazione permanente ed ereditabile del patrimonio genetico. Le mutazioni diventavano pertanto la nuova fonte di variabilità di cui la teoria dell’evoluzione per selezione naturale necessitava.
Ma la rinascita degli studi genetici che caratterizzò i primi anni del ‘900 (Hugo de Vries, Karl E. Correns, Erich von Tschermak) vide tuttavia l'aprirsi di un conflitto tra la teoria darwiniana dell'evoluzione e la teoria mendeliana dell'ereditarietà.
La caratteristica essenziale dell'ereditarietà mendeliana era infatti la possibilità di trasmettere inalterate le caratteristiche genetiche di generazione in generazione. Tutti i brillanti risultati ottenuti e la possibilità di prevedere anche quantitativamente la composizione delle generazioni filiali si fondavano infatti su questo fondamentale assunto.
Il darwinismo invece postulava la possibilità di trasformazione della specie per lento accumulo di lievi differenze nei caratteri originari. La stessa teoria ereditaria proposta da Darwin (pangenesi) prevedeva una specie di mescolamento e quindi di diluizione dei caratteri materni e paterni per fusione delle gemmule riproduttive (in questo Darwin rimaneva ancorato alla tradizione secolare degli allevatori che ammetteva che i caratteri dei figli si producessero per mescolamento del sangue dei genitori. Termini come “mezzosangue”, “purosangue” “sanguemisto” sono ancor oggi d’uso comune).
Il divario tra i due approcci si acuì ulteriormente quando si iniziarono a studiare i caratteri quantitativi a variabilità continua. Oggi noi sappiamo che anch'essi sono riconducibili ad un'eredità di tipo mendeliano con un controllo polifattoriale del carattere, ma allora l'interpretazione corrente era che si trattasse di una variabilità puramente fenotipica, non ereditabile. Sembrava dimostrarlo anche la legge della regressione filiale, tipica dei caratteri a variabilità continua, per cui i figli tendono a presentare caratteristiche di tipo medio, anche quando derivano da genitori che presentano caratteri estremi.
I biologi che si occupavano verso i primi anni del '900 dello studio dei caratteri quantitativi misurabili fondarono la biometria.
La scuola biometrica (K.Pearson, W.Weldon) partiva dall’evidenza osservativa che la variabilità era di tipo continuo nella maggior parte degli organismi. Così, mentre i biometrici continuarono ad aderire alla concezione darwiniana di evoluzione come processo lento e graduale, i mendeliani ritenevano che le piccole variazioni caratteristiche dei caratteri quantitativi, fossero in gran parte fenotipiche, non riconducibili all'eredità mendeliana, quindi non ereditabili e non invocabili, come aveva fatto Darwin, per giustificare il processo evolutivo.
Essi ritenevano che l'unica variabilità in grado di essere ereditata fosse quella prodotta in modo vistoso dalle mutazioni, trasmissibile secondo i meccanismi mendeliani, producendo un processo evolutivo non graduale e progressivo, ma a salti.
Secondo i mendeliani qualsiasi tentativo di selezione effettuato su piccole variazioni quantitative sarebbe inutile, reso inefficace dalla regressione.
Un altro elemento di incertezza all'inizio del secolo era legato al ruolo della riproduzione sessuale nei processi evolutivi. Perché la riproduzione sessuale non porta ad un aumento di uniformità, inaridendo in tal modo la fonte di variabilità necessaria al processo evolutivo?
A questo e ad altri interrogativi si iniziò a rispondere ben presto e la frattura aperta all'inizio del secolo tra la teoria "atomistica" mendeliana della genetica e la descrizione "biometrica" darwiniana di una variazione continua nelle popolazioni reali si ricompose rapidamente.
Nel 1908 l'americano Hardy ed il tedesco Weinberg dimostrarono, indipendentemente l’uno dall'altro che la riproduzione sessuale non modifica la frequenza degli alleli all'interno di una popolazione. In altre parole l'eredità mendeliana conserva la variabilità durante il ciclo riproduttivo.
Successivamente fu possibile evidenziare sperimentalmente che i caratteri a variabilità continua potevano essere interpretati in base ad un'analisi plurifattoriale di tipo mendeliano, dimostrando in tal modo che anche le piccole variazioni presenti nelle popolazione potevano avere un fondamento genetico e costituire materiale per il processo evolutivo.
La teoria sintetica dell’evoluzione
Nel 1918 Ronald A. Fisher nel suo studio intitolato "The Correlation Between Relatives on che Supposition of Mendelian Inheritance." dimostrò che tutti i risultati del processo continuo descritto da Darwin possono esser fatti derivare dai principi mendeliani. Fisher dimostrò come la variabilità continua possa essere il risultato dell’effetto additivo di molti geni. Questo lavoro è spesso considerato l’inizio della sintesi tra i due modelli, poichè Fischer aveva reso disponibile un rigoroso modello statistico-matematico in grado di soddisfare le esigenze (ed i metodi) sia della scuola biometrica che di quella mendeliana.
Fu lo stesso Fisher (The Genetical Theory of Natural Selection, 1930) e, indipendentemente, John B. S. Haldane (The Causes of Evolution, 1932) e Sewall Wright (Evolution in mendelian populations, 1931), a completare il lavoro teorico che portò alla sintesi della teoria darwiniana della selezione naturale con quella mendeliana dell'ereditarietà. Nasce il neodarwinismo, o teoria sintetica dell'evoluzione.
Si stabilisce, dunque, che la selezione naturale agisce sulla normale variabilità delle popolazioni selvatiche, che seguono le leggi di ereditarietà mendeliane. Nasce la genetica delle popolazioni. Le prime applicazioni si devono a Theodosius Dobzhansky, il quale verificò sperimentalmente, a partire dal 1935, la validità della teoria sintetica dell'evoluzione in popolazioni di Drosophila (ancor oggi uno dei modelli preferiti dai genetisti). Del 1937 è il suo fondamentale "Genetics and the Origin of Species". La moderna sintesi (come spesso è definita la teoria sintetica) finisce per influenzare e rivoluzionare molti campi della biologia. Ne dà un esempio Julian Huxley, che applica i concetti del neodarwinismo alla tassonomia Del 1940 è il suo "The New Systematics",in cui verifica la piena compatibilità della nuova teoria con le vecchie categorie sistematiche. Sulla stessa strada è il fondamentale "Systematics and the Origin of Species" (1942) di Ernst Mayr.
Contemporaneamente anche altre branche della biologia come la paleontologia, che inizialmente erano rimaste relativamente ai margini del dibattito iniziarono a reinterpretare le proprie conoscenze alla luce della nuova teoria sintetica che divenne ben presto il paradigma e la nuova struttura unificante delle diverse discipline biologiche.
Negli anni '40 la nuova teoria era ormai completa ed organicamente fondata. La sua nuova immagine interdisciplinare prese forma nel libro di Huxley"Evolution. The Modern Synthesis" (1942).
La genetica di popolazione
La genetica di popolazione studia la struttura genetica delle popolazioni (variabilità e distribuzione dei caratteri all’interno di un gruppo di individui) ed i processi evolutivi ai quali essa è sottoposta. (dinamica delle variazioni di frequenza dei caratteri e cause che le provocano).
Tale approccio è essenziale per una corretta comprensione dei processi evolutivi. L'evoluzione, a livello di base, è infatti determinata dai cambiamenti progressivi che subiscono le frequenze alleliche nelle popolazioni fino a trasformarsi in nuove specie (speciazione). La popolazione viene considerata l’unità di base del cambiamento evolutivo.
La specie è l'unità tassonomica elementare, costituita da tutti individui morfologicamente simili, tra loro fecondi ed in grado di generare prole fertile. Tale definizione è purtroppo limitata solamente a quelle specie che si riproducono per via sessuata. Nel caso di organismi che si riproducono, come ad esempio i batteri, per via asessuata, la specie deve essere individuata attraverso altri criteri (morfologici, biochimici etc).
Una specie è in genere formata da più popolazioni.
Una popolazione è costituita da un gruppo di individui della stessa specie che, vivendo in un'area geografica particolare, si è ad essa adattata e presenta pertanto una struttura genetica diversa da quella di un’altra popolazione. Gli individui di una popolazione, presentando una maggior probabilità di incrocio reciproco rispetto agli individui di un'altra popolazione, tendono a mantenere nel tempo le differenze genetiche rispetto alle altre popolazioni.
Una popolazione è univocamente descritta dal suo pool genico, inteso come l'insieme di tutti gli alleli presenti nella popolazione, descritti per tipo e frequenza. La struttura genetica di una popolazione viene espressa in termini di frequenze alleliche e, di conseguenza, genotipiche.
Il pool genico è ovviamente un concetto astratto, essendo in pratica distribuito all’interno degli individui che compongono la popolazione. Il singolo individuo è solo un temporaneo contenitore di un piccolo campione del pool genico.
La genetica di popolazione studia la struttura ed i cambiamenti di composizione del pool genico (sia come tipo che come frequenza degli alleli) e le forze che li provocano (fattori evolutivi).
Nelle popolazioni naturali alcuni alleli aumentano di frequenza da una generazione ad un'altra ed altri diminuiscono, alcuni spariscono ed altri nuovi si producono.
Se un individuo presenta una combinazione favorevole di alleli nel suo genotipo, questi alleli hanno una probabilità maggiore di essere presenti in proporzione maggiore nel pool genico della generazione successiva. Se la combinazione non è favorevole la frequenza d quegli alleli nella generazione successiva diminuisce o, allimite, si annulla.
L'evoluzione è il risultato di tali cambiamenti accumulati nel pool genico col passar de tempo.
Lo strumento statistico-matematico utilizzato per fotografare il pool genico è fornito dalla cosiddetta legge di Hardy-Weinberg che correla le frequenze genotipiche di un carattere in una popolazione alle frequenze alleliche del medesimo carattere nel pool genico.
Come vedremo tale relazione permette di calcolare le frequenze alleliche, note le frequenze genotipiche. Infatti, poiché il pool genico è un’entità astratta, le frequenze alleliche non sono direttamente misurabili, mentre le frequenze dei diversi genotipi in una popolazione in alcuni casi lo sono.
La legge di Hardy-Weinberg è applicabile solo se la popolazione è in equilibrio genetico, detto anche equilibrio di Hardy-Weinberg.
Si dice che una popolazione è in equilibrio genetico quando tutte le frequenze alleliche rimangono costanti nel tempo. In altre parole quando non è in atto nessun processo evolutivo.
Affinché in una popolazione valga la legge di Hardy-Weinberg è necessario che siano soddisfatte le seguenti condizioni:
- la popolazione deve essere sufficientemente numerosa in modo che siano applicabili le leggi della probabilità (in questo modo le frequenze che misuriamo sono una buona stima della probabilità)
- l'accoppiamento tra gli individui della popolazione deve essere assolutamente casuale (panmissia).
- Tutti gli individui si accoppiano e si riproducono (non vi è selezione naturale)
- Non devono essere presenti altri fattori in grado di far variare le frequenze alleliche, come mutazioni, movimenti di individui tra popolazioni (flusso genico) e deriva genetica.
Supponiamo ora che un carattere sia presente nella popolazione con 2 alleli (A, a ) indichiamo la frequenza dell'allele dominante nel pool genico con 'p' e quella dell'allele recessivo 'q'. Nel caso di incroci perfettamente casuali sarà allora possibile calcolare le frequenze con cui compaiono i 3 genotipi (AA, Aa, aa) nella popolazione con un quadrato di Punnett.

Per ottenere le frequenze genotipiche è quindi sufficiente sviluppare il quadrato della somma delle frequenze alleliche. Otteniamo così la formula di Hardy-Weinberg
(p + q)2 = p2 + 2pq + q2
Dove
p = frequenza allele dominante (A) nel pool genico
q = frequenza allele recessivo (a) nel pool genico
p2 = frequenza degli omozigoti dominanti (AA) nella popolazione
2pq = frequenza degli eterozigoti (Aa) nella popolazione
q2 = frequenza degli omozigoti recessivi (aa) nella popolazione
Si noti che la somma delle frequenze alleliche (p + q) deve sempre essere uguale ad 1.
Nel caso di un carattere presente nel pool genico con più di due alleli si dovrà comunque sviluppare il quadrato della somma di tutti gli alleli.
Ad esempio nel caso dei gruppi sanguigni sistema AB0 con i 3 alleli A, B e 0 avremo
p = frequenza allele A nel pool genico
q = frequenza allele B nel pool genico
r = frequenza allele 0 nel pool genico
(p + q + r)2 = p2 + q2 + r2 + 2pq + 2pr + 2qr
dove
p2 = frequenza individui AA nella popolazione
q2 = frequenza individui BB nella popolazione
r2 = frequenza individui 00 nella popolazione
2pq = frequenza individui AB nella popolazione
2pr = frequenza individui A0 nella popolazione
2qr = frequenza individui B0 nella popolazione
Se la popolazione è in equilibrio genetico si può utilizzare la legge di Hardy-Weinberg per calcolare le frequenze alleliche misurando le frequenze degli omozigoti recessivi nella popolazione. Si noti infatti che gli omozigoti recessivi sono in genere gli unici di cui si possa misurare la frequenza nella popolazione poiché il loro fenotipo è distinguibile da quello degli omozigoti dominanti e degli eterozigoti, che invece presentano il medesimo fenotipo.
Un esempio aiuterà a chiarire.
La fenilchetonuria o PKU (o iperfenilalaninemia) è una grave malattia genetica, in cui gli omozigoti recessivi mancano di un enzima in grado di utilizzare correttamente l'aminoacido fenilalanina, normalmente presente nei cibi, con conseguente ritardo mentale dovuto all'azione tossica della fenilalanina e dei suoi derivati sulle cellule cerebrali.
I dati statistici dicono che un neonato su 15.000 è affetto da questa malattia.
Se dunque la frequenza degli omozigoti recessivi q2 = 1/15.000, per calcolare la frequenza dell'allele recessivo nel pool genico della popolazione sarà sufficiente estrarre la radice quadrata di 1/15.000


q = 0.008
p = 1 - q = 0.992
GLi omozigoti dominanti saranno
p2 =0.984 = 98,4%
mentre gli eterozigoti, portatori sani, saranno 2pq = 0.016 = 1,6%
cioè circa 16 persone su 1000.
Nel caso di alleli multipli la frequenza di ciascun allele è pari alla frequenza del suo omozigote più la semisomma delle frequenze di tutti gli eterozigoti in cui l’allele compare. Verifichiamolo, ad esempio nel caso di un carattere che si presenta con tre alleli (A, B e C) aventi rispettivamente frequenze p, q ed r. Calcoliamo la frequenza r dell’allele C nel pool genico. Ricordando che (p + q + r) = 1, potremo scrivere
r = r(p + q + r) = pr + qr + r2 = (2pr + 2qr)/2 + r2
dove
(2pr + 2qr)/2 = semisomma delle frequenze degli eterozigoti in cui compare l’allele C
r2 = frequenza degli omozigoti CC
Come si ricorderà, storicamente la legge di Hardy-Weinberg ha una sua rilevanza in quanto dimostrò che la riproduzione sessuata non distruggeva la variabilità genetica all’interno di una popolazione, come poteva a prima vista sembrare.
Vediamo un esempio. Si consideri una popolazione costituita da:
36% di omozigoti dominanti AA
48% di eterozigoti Aa
16% di omozigoti recessivi aa
Le frequenze alleliche nel pool genico di questa popolazione si possono calcolare a partire dalla percentuale di omozigoti recessivi.
La frequenza q dell’allele recessivo a è pari a

La frequenza p dell’allele dominante A è pari a
P = 1 – q = 1 – 0,4 = 0,6
Ora, se gli individui di questa popolazione si incrociano casualmente avremo che
Gli individui omozigoti dominanti produrranno solo gameti con l’allele A, gli individui eterozigoti produrranno metà gameti con l’allele A e metà gameti con l’allele a, gli individui omozigoti recessivi produrranno solo gameti con l’allele recessivo, nelle seguenti proporzioni
individui |
gameti |
Totale gameti |
Omozigoti dominanti AA |
36% gameti A |
60% gameti A |
Eterozigoti Aa |
24% gameti A 24% gameti a |
|
|
|
40% gameti a |
Omozigoti recessivi aa |
16% gameti a |
Se durante la riproduzione i gameti si uniscono casualmente, otterremo

Come si può osservare le frequenze genotipiche sono rimaste inalterate e di conseguenza anche le frequenze alleliche
 In realtà le condizioni poste dalla legge di Hardy-Weinberg sono estremamente restrittive e molto difficilmente possono essere soddisfatte nelle popolazioni naturali. La legge di Hardy-Weinberg fornisce pertanto un quadro teorico di riferimento rispetto al quale confrontare le popolazioni reali.
In realtà le condizioni poste dalla legge di Hardy-Weinberg sono estremamente restrittive e molto difficilmente possono essere soddisfatte nelle popolazioni naturali. La legge di Hardy-Weinberg fornisce pertanto un quadro teorico di riferimento rispetto al quale confrontare le popolazioni reali.
Microevoluzione e fattori evolutivi
Le popolazioni evolvono, trasformando il loro pool genico nel tempo. In tal caso dobbiamo ricercare i fattori che modificano le frequenze alleliche. I fattori che modificano le frequenze alleliche, avendo sul pool genico effetti direttamente osservabili e misurabili, sono detti fattori evolutivi e sono alla base della cosiddetta microevoluzione.
I fattori evolutivi sono 5: le mutazioni, le modalità di incrocio, la deriva genetica, il flusso genico e la selezione naturale.
Mutazioni
Le mutazioni sono alterazioni casuali ed ereditarie del corredo cromosomico, che comportano la modificazione o la comparsa di nuovi alleli.
Come abbiamo già visto le mutazioni possono essere indotte da agenti mutageni, ma esistono anche mutazioni cosiddette "spontanee" che avvengono senza che sia possibile evidenziare gli agenti fisici o chimici che le hanno indotte.
La frequenza delle mutazioni spontanee è in genere bassa. Per mutazioni osservabili sul fenotipo dell'uomo essa varia da 1 su 1000 fino a 1 su 10.000 gameti per generazione, a seconda dell'allele interessato.
Si è calcolato che ogni nuovo essere umano è mediamente portatore di due nuove mutazioni. Perciò, anche se l'incidenza delle mutazioni per individuo è molto bassa, il numero di nuove mutazioni per generazione nel pool genico è piuttosto elevata.
Flusso genico
Il flusso genico è connesso con il processo di migrazione di individui riproduttivi tra popolazioni della stessa specie o, come nel caso delle piante e degli invertebrati acquatici, con il movimento del polline o dei gameti liberati nell'acqua. Il flusso genico può portare nuovi alleli all'interno di una popolazione o può mutare la frequenza allelica.
Deriva genetica
La deriva genetica o genetic drift è una variazione della frequenza allelica dovuta al caso. Si hanno fenomeni di deriva genetica quando da una popolazione viene estratto e separato un piccolo gruppo di individui che va a formare una nuova popolazione. Il fenomeno è legato al fatto che più piccolo è un campione e maggiori sono le probabilità che esso risenta di fluttuazioni statistiche casuali e non sia pertanto rappresentativo della popolazione di origine. Da un punto di vista statistico si tratta dunque di un errore di campionamento. Le conseguenze della deriva genetica sono un cambiamento casuale delle frequenze alleliche ed una riduzione della variabilità genetica.
Vi sono due casi tipici di deriva genetica descritti dai genetisti come effetto del fondatore ed effetto collo di bottiglia,
L'effetto del fondatore si produce quando un piccolo numero di individui si separa da una popolazione per andare a formare una nuova colonia. In questo caso è poco probabile che il pool genico di questo piccolo gruppo presenti gli stessi alleli e con la stessa frequenza del pool genico della popolazione madre. In altre parole esso non è un campione statisticamente rappresentativo. Un po' come è assai improbabile che su dieci lanci di una moneta otteniamo sempre cinque volte testa e cinque volte croce.
Un classico esempio dell'effetto del fondatore è quello della comunità Amish in Pennsylvania. Si tratta dei discendenti di un piccolo gruppo di individui immigrati dalla Svizzera durante il 1700. Uno dei fondatori possedeva casualmente un allele recessivo che determina una rara forma di nanismo e polidattilia. Attualmente il 13% degli individui della colonia reca questo allele, con una frequenza enormemente superiore a quella riscontrabile nella popolazione mondiale.
L’effetto collo di bottiglia si produce quando una popolazione viene decimata da un evento catastrofico, come un'epidemia, una eccessiva predazione, un'eruzione vulcanica etc. Quando una popolazione esce da un collo di bottiglia è molto probabile che il suo pool genico sia diverso da quello originario e sicuramente con una variabilità inferiore.

Una perdita di variabilità genica è stata documentata nell’elefante marino boreale e nel ghepardo. Nell’Ottocento l’elefante marino fu cacciato fin quasi all’estinzione, tanto che alla fine di quel secolo ne sopravvivevano solo circa 20 esemplari. Grazie a una politica di protezione, la popolazione è oggi cresciuta a circa 30000 individui, ma le analisi biochimiche mostrano che tutti gli elefanti marini sono quasi identici dal punto di vista genico. A causa della scarsa variabilità genica, questa specie ha quindi una potenzialità di evolversi in risposta alle variazioni ambientali notevolmente ridotta e, in definitiva, benché il numero di individui si sia accresciuto, la specie nel suo complesso è ancora da ritenersi in pericolo di estinzione. I ghepardi presentano lo stesso problema, benché si ignori a che cosa sia dovuto il collo di bottiglia. Quindi, anche i ghepardi potrebbero essere in grande pericolo nel caso intervengano piccoli cambiamenti ambientali.
Modalità di incrocio
Abbiamo visto come la legge di Hardy-Weinberg dimostri che quando in una popolazione l'accoppiamento tra individui è assolutamente casuale (panmissia), le frequenze alleliche e genotipiche tendono a rimanere costanti nel tempo, poiché vi è la massima probabilità che ogni allele si distribuisca uniformemente nel pool genico.
Il termine “accoppiamento casuale” significa che gli individui non scelgono il loro compagno sulla base di un qualche carattere ereditabile. Ad esempio, gli esseri umani si accoppiano casualmente rispetto ai gruppi sanguigni poiché in genere essi non scelgono il loro compagno sulla base di questa caratteristica.
Tuttavia è necessario tener presente che in genere all’interno di una specie sono presenti dei sottogruppi genetici (razze, etnie etc) il cui pool genico differisce e spesso gli individui tendono ad incrociarsi all’interno del proprio sottogruppo (endogamia). In questo senso, se gli italiani ed i russi mostrano frequenze diverse per il gruppo sanguigno AB0, e gli italiani tendono ad accoppiarsi con gli italiani, mentre i russi si accoppiano con i russi, esisterà un involontario accoppiamento non casuale rispetto al gruppo sanguigno AB0.
Tutti i sistemi di incrocio che comportano una deviazione dalla casualità (ad esempio la scelta del partner in base al fenotipo o selezione sessuale, tendenza all'incrocio tra consanguinei etc) portano ad una modificazione delle frequenze genotipiche e possono modificare il pool genico.
Esempi di incroci non casuali sono l’inincrocio e l’incrocio assortativo.
La modalità di incrocio che si ha quando determinati genotipi sono responsabili di caratteristiche fenotipiche che influenzano la scelta del partner, è detta accoppiamento assortativo positivo. Si ha invece accoppiamento assortativo negativo quando l’accoppiamento risulta preferenziale tra individui aventi differenti genotipi. La selezione sessuale è una forma di selezione naturale come conseguenza di un accoppiamento assortativo in cui gli individui scelgono il partner non in modo casuale, ma in relazione a particolari caratteristiche, indipendentemente dagli effetti che tali caratteristiche potrebbero avere sulla capacità di sopravvivenza dell'individuo.
Come lo stesso Darwin aveva compreso, molti dei più appariscenti adattamenti degli animali non hanno direttamente a che fare con la sopravvivenza, ma con la selezione sessuale. La selezione sessuale è la causa principale del dimorfismo sessuale.
Nelle specie in cui sono le femmine che scelgono il partner per l'accoppiamento (e sono le più numerose), i maschi tendono ad essere molto più vistosi e colorati, proprio perché tali caratteri sono stati preferiti e quindi selezionati dalle femmine della propria specie.
L'inincrocio o inbreeding è l'accoppiamento sistematico tra individui strettamente imparentati (consanguinei). Si ha quando l’accoppiamento fra individui imparentati è più frequente rispetto a quanto atteso in base al caso. In alcune piante ed animali ermafroditi l'inincrocio può giungere al caso estremo dell'autofecondazione.
Se tale pratica si mantiene per più generazioni produce una modificazione delle frequenze genotipiche con una tendenza all'eliminazione degli individui eterozigoti e all'affermazione dell'omozigosi (line pure).
Se ad esempio consideriamo una popolazione costituita da 2000 individui eterozigoti Aa, dopo quattro generazioni di inincrocio si assiste ad una modificazione delle frequenze genotipiche con la presenza di un 87,5% di omozigoti ed un 12,5% di eterozigoti.

Ad ogni generazione si dimezza il numero degli eterozigoti. Infatti quando gli eterozigoti si incrociano tra loro producono un 50% di eterozigoti, un 25% di omozigoti dominanti ed un 25% di omozigoti recessivi (legge della segregazione), mentre quando gli omozigoti si incrociano tra loro producono ovviamente il medesimo genotipo.
In effetti la frequenza allelica non è mutata, infatti l'allele 'A' e l'allele 'a' presentano ciascuno una frequenza del 50%, invariata rispetto alla popolazione iniziale.
Tuttavia, poiché molti caratteri letali o che esprimono difetti o malattie sono recessivi, l'inincrocio tende a portarli alla luce allo state omozigote, esponendoli alla selezione naturale e favorendone l'eliminazione. Alleli recessivi indesiderati e rari, che in condizioni normali sono presenti prevalentemente allo stato eterozigote e che quindi risultano latenti, hanno ora una maggiore probabilità di esprimersi allo stato omozigote.
Negli esseri umani ad esempio è noto che rare anomalie recessive si manifestano più frequentemente nella prole di genitori consanguinei.
E' stato inoltre osservato che una maggior percentuale di omozigosi è spesso associata (soprattutto negli animali) a manifestazioni di scarsa vitalità e di debolezza costituzionale, a prescindere dalla presenza di caratteri nocivi di tipo recessivo (depressione da inincrocio).
Il fenomeno è probabilmente da mettere in relazione con il fatto che la condizione eterozigote, essendo portatrice di una maggior variabilità (maggiore è il tasso di eterozigosi in un individuo e più elevato è il numero di alleli diversi che possiede) potrebbe essere favorita dalla selezione naturale. Tale aspetto è anche noto come superiorità dell’eterozigote e risulta particolarmente evidente nel fenomeno dell’eterosi (o lussureggiamento degli ibridi o vigore ibrido).
L’eterosi è un fenomeno opposto e complementare a quello della depressione da inbreeding. Se l’inbreeding è il risultato dell’accoppiamento di individui simili (parenti) e in dipendenza di ciò produce un aumento di omozigosi a scapito dell’eterozigosi, l’eterosi è il risultato dell’aumento di eterozigosi come conseguenza dell’accoppiamento tra individui diversi, provenienti, ad esempio, da razze o da linee pure differenti.
Se si incrociano infatti due linee pure diverse in molti casi si ottengono individui con una altissima percentuale di eterozigosi (ibridi), che presentano una straordinaria vigoria rispetto alle linee parentali. II fenomeno è stato sfruttato su larga scala per la produzione di ibridi di mais.
Altri fattori legati ai processi di riproduzione che favoriscono la variabilità all'interno delle popolazioni sono la diploidia e la sessualità.
Nella sessualità comprendiamo qualsiasi processo attraverso il quale gli individui scambiano materiale genetico al fine di aumentare la loro variabilità o quella della loro discendenza. II modo di gran lunga più diffuso attraverso cui ciò avviene è mediante la gametogenesi e la successiva fecondazione.
Questi processi producono nuove combinazioni geniche, aumentando enormemente la variabilità all'interno delta popolazione, in tre modi
- crossing-over e relativa ricombinazione genica
- assortimento indipendente dei cromosomi
- unione di genomi parentali differenti durante la fecondazione
Anche la condizione diploide infine può essere considerata un elemento che favorisce la variabilità all'interno della popolazione.
In un organismo aploide infatti tutte le variazioni genetiche sono espresse immediatamente nel fenotipo e sono quindi esposte al processo di selezione. In un organismo diploide invece tali variazioni possono conservarsi allo stato recessivo e andare così ad aumentare la variabilità senza subire alcuna pressione selettiva.
Selezione naturale e fitness
Abbiamo già visto come la teoria darwiniana dell’evoluzione debba essenzialmente giustificare due fenomeni: l’origine della diversità degli esseri viventi e l’origine dell’adattamento degli stessi all’ambiente. La risposta di Darwin a questo secondo punto è che in un ambiente in cui le risorse non sono sufficienti si instaura una lotta per la sopravvivenza. Il risultato di questa competizione è che gli individui che possiedono la combinazione genica più efficiente relativamente ad un certo ambiente (cioè sono più adatti) hanno maggiori probabilità di sopravvivere e di riprodursi, trasmettendo le loro caratteristiche genetiche alle generazioni successive. Questo meccanismo è chiamato da Darwin selezione naturale e porta ad un progressivo adattamento della popolazione al suo ambiente.
La capacità di sopravvivere fino a riprodursi è dunque un aspetto fondamentale, tanto che oggi si preferisce descrivere la selezione naturale come una differenza nel tasso di riproduzione (differente successo riproduttivo)degli organismi di una popolazione in un determinato ambiente, piuttosto che come una lotta per la sopravvivenza.
La probabilità che un genotipo ha di sopravvivere e di riprodursi relativamente ad un certo ambiente, viene misurata ed è chiamata fitness darwiniana (w). La fitness è una misura dell’efficienza riproduttiva di un genotipo.
Un medesimo genotipo potrà avere fitness diverse in ambienti diversi.
La fitness darwiniana (o riproduttiva) non va confusa con la “fitness fisica”. Un individuo può essere forte, sano ed intelligente, ma se è sterile la sua fitness riproduttiva è nulla.
Così un carattere che raddoppia la fecondità di un individuo e contemporaneamente ne dimezza la vita, comunque ne aumenta la fitness.
La fitness si misura in termini relativi di un genotipo rispetto ad un altro. Il genotipo che ha la maggior probabilità di sopravvivenza riproduttiva ha fitness = 1, gli altri presentano fitness inferiori all’unità.
Ad esempio possiamo calcolare la fitness relativa dei genotipi AA, Aa ed aa sulla base della loro sopravvivenza o fecondità differenziale nell’arco di una generazione (dati demografici)
Genotipi |
AA |
Aa |
aa |
totale |
Numero di individui parentali |
40 |
50 |
10 |
100 |
Numero di individui prodotti in F1 da ciascun genotipo |
80 |
90 |
10 |
180 |
Numero medio di individui generati da ciascun genotipo in F1 |
80/40=2 |
90/50=1,8 |
10/10=1 |
|
Fitness |
2/2 = 1 |
1,8/2 = 0,9 |
1/2 = 0,5 |
|
Una misura correlata alla fitness è il coefficiente di selezione (s), definito come s = 1 – w.
Il coefficiente di selezione misura la riduzione di fitness per un dato genotipo rispetto a quello con fitness massima (per geni letali w = 0 ed s = 1).
Nell’esempio precedente il genotipo Aa presenta fitness = 0,9 (probabilità di sopravvivenza e riproduzione pari al 90% rispetto al genotipo AA con fitness massima) e quindi un coefficiente di selezione = 0.1 (probabilità di sopravvivenza e riproduzione del 10% in meno rispetto al genotipo AA con fitness massima).
Da un punto di vista molto generale si possono distinguere tre modalità attraverso le quali agisce la selezione naturale: selezione direzionale, divergente e stabilizzante.
Ciascuno di questi tre tipi di selezione agisce modificando la distribuzione di frequenza (gaussiana) di un determinato carattere all’interno di una popolazione.
Si parla di selezione direzionale quando la selezione naturale favorisce gli individui che si trovano ad uno dei due estremi dell’intervallo di variabilità del carattere (individui rari). Questi individui che prima erano poco frequenti nella popolazione, iniziano a godere di un vantaggio adattativo e diventano più numerosi. L’effetto è uno spostamento laterale della gaussiana nella direzione degli individui premiati dalla selezione naturale.

Un esempio classico di selezione direzionale è quello avvenuto a carico di Biston betularia, una farfalla che presenta un allele dominante per il colore nero (forma melanica) ed uno recessivo per il colore chiaro. Prima dell'avvento della rivoluzione industriale in Gran Bretagna le farfalle chiare erano favorite poiché si mimetizzavano più facilmente sui tronchi degli alberi (betulle) e sui muri, evitando di essere predate dagli uccelli.
Quando nella seconda metà dell’Ottocento la fuliggine e lo smog che accompagnarono l'industrializzazione ebbero annerito gran parte degli alberi e delle case, le farfalle chiare divennero facile preda, mentre le farfalle scure vennero favorite essendo più adatte al nuovo ambiente (melanismo industriale).
Esempi di selezione direzionale sono anche la comparsa della resistenza agli antibiotici in certi batteri, o agli insetticidi e agli anticrittogamici nei parassiti. In realtà non sarebbe dunque corretto parlare di una “comparsa”. Gli alleli responsabili della resistenza sono già presenti in alcuni individui che però sono inizialmente in numero esiguo nella popolazione. Quando la popolazione viene trattata (con antibiotici, antiparassitari etc), vengono in pratica selezionati solo gli individui già geneticamente resistenti. Questi sono gli unici che riescono a riprodursi e diverranno preponderanti nelle generazioni successive.
Quando la selezione naturale favorisce entrambe le forme estreme dell’intervallo di variabilità si parla di selezione divergente. La gaussiana tende ad aprirsi formando due distribuzioni a campana sovrapposte.

Si parla infine di selezione stabilizzante quando vengono favoriti gli individui intermedi. In questo caso la gaussiana tende ad alzarsi e a restringersi intorno alla media.

Il numero delle uova deposte dagli uccelli è un esempio di selezione stabilizzante. In genere il numero di uova deposte è determinato geneticamente in ogni specie. Si è potuto osservare sperimentalmente che gli uccelli che depongono un numero di uova conforme alla media della loro specie, hanno un maggior numero di nati che sopravvivono, rispetto agli individui della stessa specie che producono un numero maggiore a minore di uova.
Sorprendentemente ciò accade anche negli esseri umani dove attualmente la media del peso corporeo alla nascita e di 3.2 kg. ed i neonati più grossi e quelli più piccoli hanno una minor probabilità di sopravvivenza rispetto a quelli con peso medio.
La selezione stabilizzante viene a volte confusa con la selezione bilanciata.
La selezione bilanciata è una particolare forma di selezione naturale che favorisce il mantenimento di diversi alleli di uno stesso gene all'interno di una popolazione. Se ne ha un esempio nei casi di vantaggio eterozigote (o sovradominanza) e di selezione dipendente dalla frequenza.
Il vantaggio eterozigote si manifesta quando un individuo eterozigote per un particolare locus è più adatto alla sopravvivenza di un individuo omozigote. È questo il caso ad esempio della drepanocitosi (anemia falciforme). Gli individui che sono omozigoti per l'allele recessivo, presentano l’emoglobina mutata (emoglobina S) e risultano affetti dal disturbo (globuli rossi a forma di falce). Al contrario, un individuo eterozigote sarà caratterizzato solo da lievi imperfezioni nella forma delle cellule sanguigne, che le rendono tuttavia resistenti alla malaria. Questa forma di resistenza è favorita dalla selezione naturale nelle regioni tropicali, dove la diffusione della malaria (causata dal parassita Plasmodium falciparum) rende la situazione eterozigote molto favorevole dal punto di vista evolutivo. In questo modo le frequenze dei fenotipi omozigoti ed eterozigoti si mantengono relativamente stabili.
Si parla in particolare di polimorfismo bilanciato quando la selezione bilanciata che agisce all'interno di una popolazione è in grado di mantenere una frequenza relativamente stabile di due o più forme fenotipiche.
Speciazione
L’insieme dei fenomeni che portano alla nascita di una nuova specie è detto speciazione.
La speciazione può essere filetica o divergente.
La speciazione filetica (o anagenesi) agisce in modo lineare attraverso la lenta, trasformazione di una specie in un'altra che sostituisce la prima. In questo caso la specie si adatta all’ambiente che cambia gradualmente e si trasforma anch’essa di conseguenza. Se questo fosse stato l’unico tipo di speciazione ovviamente oggi la terra sarebbe popolata da un’unica specie. Più interessante per i biologi è invece la speciazione divergente in cui una specie genera due (cladogenesi) o più (radiazione adattativa) linee filetiche che evolvono in altrettante specie.
La radiazione adattativa è uno del fenomeni evolutivi meglio documentati dai fossili. Secondo alcuni autori essa rappresenta la principale modalità di speciazione. Si presenta nei reperti fossili come una vera e propria esplosione evolutiva, con l'improvvisa comparsa di numerose nuove specie aventi tutte un antenato in comune.
La radiazione adattativa è in genere associata all'apertura improvvisa di una nuova frontiera biologica, dove una specie viene ad avere a disposizione numerose e diversificate nicchie ecologiche da colonizzare.
II termine nicchia ecologica designa l'insieme delle attività e dei rapporti che ogni specie intrattiene con il suo habitat e le particolari specializzazioni adottate per sfruttarlo. In questo modo mentre possono coesistere più specie in uno spesso habitat, due specie non possono, per definizione, occupare la stessa nicchia ecologica.
Così, ad esempio, nelle grandi praterie dell'Africa convivono molti mammiferi erbivori, i quali non entrano però in competizione per il cibo, occupando nicchie diverse. Le zebre si cibano della parte superiore delle erbe alte, gli gnu della parte media, le gazzelle delle erbe basse e dei frutti caduti.
La radiazione adattativa si può verificare quando
- sono rese disponibili nuove nicchie, non precedentemente occupate. È il caso della colonizzazione da parte delle prime piante e dei primi animali della terraferma.
- si liberano improvvisamente nicchie precedentemente occupate. L'esempio più eclatante si ha con la radiazione dei mammiferi, i quali, dopo aver coabitato per milioni di anni all'ombra dei grandi rettili, sono letteralmente esplosi al momento della loro estinzione, occupando molte delle nicchie liberatesi.
- le nuove specie risultano più adatte ad occupare vecchie nicchie e si sostituiscono a chi le occupa. Un esempio si è avuto con la sostituzione in sudamerica dei carnivori marsupiali indigeni con i carnivori placentati provenienti dal nord, probabilmente migliori predatori.
La speciazione divergente avviene quando alcune popolazioni che costituiscono una specie si trasformano in nuove specie.
Ricordiamo che la specie è definita come un insieme di individui interfecondi ed i cui membri non possono avere incroci fertili con gli individui appartenenti ad un'altra specie. Dunque la condizione indispensabile affinché una o più popolazioni all'interno di una specie si separino fine a diventare esse stesse nuove specie, è che tra le due popolazioni vi sia un'interruzione del flusso genico. In altre parole se tra due popolazioni appartenenti ad una stessa specie, si produce un isolamento riproduttivo che impedisca ai loro membri di scambiarsi geni, le eventuali differenze tra i due pool genici possono mantenersi ed eventualmente approfondirsi fino a rendere i due pool genici incompatibili al punto da formare due nuove specie.
L’isolamento riproduttivo viene dunque considerato dai biologi la condizione indispensabile affinché avvenga il processo di speciazione. Si ritiene che nella maggior parte dei casi l’isolamento riproduttivo sia una diretta conseguenza dell'instaurarsi di un isolamento geografico tra popolazioni di una stessa specie. Questo processo viene definite speciazione allopatrica (altra patria).
In particolari circostanze l'isolamento riproduttivo può manifestarsi anche senza un isolamento geografico ed in tal caso si parla di speciazione simpatrica (stessa patria).
Nella speciazione allopatrica le nuove specie si formano come conseguenza di un processo di diversificazione tra due o più popolazioni appartenenti ad un'unica specie, le quali si adattano ad ambienti diversi.
Molto spesso la diversificazione di una specie in diverse popolazioni segue una distribuzione geografica. Le variazioni fenotipiche sono cioè correlate alle graduali modificazioni di temperatura, umidità, o di qualche altra condizione ambientale.
Tale variazione graduale in un carattere o in un gruppo di caratteri è detta cline, Molte specie presentano caratteristici clini nord-sud. 0gni popolazione distinguibile all'interno di un cline prende il nome di ecotipo. E' stato possibile dimostrare che le differenze tra ecotipi sono in parte genetiche e quindi utilizzabili dalla selezione naturale per il processo evolutivo.
Le barriere geografiche possono naturalmente essere di moltissimi tipi. I luoghi in cui tipicamente si sviluppano nuove specie sono ad esempio le isole. La separazione dell'Australia dal resto dei continenti ha prodotto al suo interno un indirizzo evolutivo particolare a diverso rispetto al resto del mondo. A tutt'oggi sono stati studiati centinaia di casi di speciazione allopatrica nelle isole, soprattutto in quelle dell'oceano Pacifico (Samoa, Tahiti, Marchesi, Hawai, Galapagos, etc.).
Esempi divenuti ormai classici, riportati dallo stesso Darwin, sono quelli che si riferiscono alle diverse specie di fringuelli e di tartarughe che vivono sulle isole Galapagos. Le 14 specie di fringuelli che si trovano attualmente su queste isole si sarebbero originate da una popolazione di fringuelli terricoli provenienti dal vicino continente i quali, separati geograficamente sulle diverse isole, avrebbero dato l'avvio ad un intenso processo di speciazione allopatrica.
Le popolazioni di molti organismi, tuttavia, possono venire separate da barriere meno drastiche degli oceani. Per una pianta può diventare un'isola la vetta di una montagna, e per un pesce un piccolo stagno.
Una volta separata la popolazione isolata, che probabilmente sarà già geneticamente diversa per effetti di deriva genetica, può iniziare a divergere ulteriormente, sotto la pressione delle diverse forze selettive, adattandosi al nuovo ambiente.
Quando le differenze tra il pool genico delle due popolazioni si è modificato al punto tale che non è possibile alcun incrocio fertile anche se le due popolazioni vengono riportate in contatto, il processo di speciazione può dirsi concluso.
I casi di speciazione simpatrica sono ritenuti meno probabili e quindi meno frequenti nella storia evolutiva.
La speciazione simpatrica si può produrre quando insorgono meccanismi di isolamento riproduttivo, all'interno di una popolazione, senza che vi sia un isolamento geografico, per cui una parte della popolazione comincia a presentare un modo di vita differente rispetto al resto.
Meccanismi di questo tipo si dividono in due gruppi. I meccanismi di isolamento riproduttivo che agiscono prima dell’incrocio, impedendolo, sono detti meccanismi precopula o prezigotici. I meccanismi di isolamento riproduttivo che agiscono dopo l’accoppiamento sono detti meccanismi post-copula o post-zigotici.
Sono esempi di meccanismi prezigotici i meccanismi etologici di isolamento riproduttivo in cui gli individui presentano diversi comportamenti sessuali (periodi diversi di accoppiamento, diversi segnali di corteggiamento etc)
Vengono considerati meccanismi post-zigotici tutti i meccanismi fisiologici di isolamento come l'incompatibilità degli organi riproduttivi, l'incompatibilità dei gameti, la produzione di uno zigote non in grado di svilupparsi, ma anche la generazione di una prole non in grado di raggiungere la maturità sessuale o non fertile (ibridi sterili).
Un interessante esempio di speciazione simpatrica viene proprio dagli ibridi e dalla poliploidia.
Con il termine 'ibrido' si intende qui individui provenienti dall'incrocio di specie differenti. Gli ibridi sono quasi sempre sterili poiché la serie di cromosomi di provenienza paterna non trova nella serie materna i propri omologhi, e non sono quindi in grado di appaiarsi durante la meiosi che non risulta per questo motivo possibile.
Gli ibridi si possono generare tra gli animali, ma sono molto più frequenti nelle piante. La gramigna ad esempio è un'erba che forma con facilità ibridi con molte piante affini. Tali ibridi, che si diffondono in maniera agamica (asessuata), mediante rizomi, sono a volte molto meglio adattati rispetto alle piante originarie.
Nelle piante spesso accade che un ibrido sterile sia in grado di produrre un individuo fertile per poliploidia. La poliploidia si ha quando il corredo cromosomico dell'individuo presenta un numero di serie di cromosomi omologhi superiore a due, tre negli individui triploidi 3n, quattro nei tetraploidi 4n, e cosi' via.
La poliploidia è un evento raro negli animali, dove si riscontra per lo più in alcuni crostacei e coleotteri, mentre è molto diffusa nelle piante.
La poliploidia si origina per un errore durante la mitosi, con i cromosomi che si dividono, mentre la cellula non entra in cidodieresi.. Sperimentalmente si può ottenere tale risultato con l'impiego della colchicina, un alcaloide che blocca la migrazione dei cromosomi ai due poli della cellula.
Se la poliploidia si verifica in un ibrido sterile, ogni cromosoma sarà presente due volte, i cromosomi duplicati potranno comportarsi come due omologhi, la meiosi diverrà possibile ed ii nuovo individuo sarà fertile.
Oggi si ritiene che circa la metà delle piante con fiori abbia avuto un'origine poliploide. Un'importante esempio è dato dal frumento attuale, una pianta esaploide che si ritiene abbia avuto origine dalla combinazione di tre antenati diploidi diversi.
Molto spesso la poliploidia è accompagnata da un aumento delle dimensioni della pianta, caratteristica spesso utilizzata per produrre piante con frutti particolarmente grandi.
Un altro esempio di speciazione simpatrica si ha nelle specie parassite che riconoscono il loro bersaglio grazie alla presenza di alcuni geni che permettono loro di individuare chimicamente l'ospite da parassitare. In alcune specie si è potuto osservare la comparsa, per mutazione dei geni che codificano le proteine identificatrici, di individui in grado di riconoscere un ospite diverso, creando improvvisamente un isolamento riproduttivo rispetto agli individui non mutati.
Coevoluzione, convergenza evolutiva e preadattamento
Abbiamo già avuto modo di sottolineare il fatto che l'evoluzione rappresenta un progressivo fenomeno di adattamento delle specie alle diverse condizioni ambientali. Qui vogliamo dare alcuni esempi particolarmente significativi di adattamento.
Si configura un processo di coevoluzione, quando due o più specie interagiscono tanto strettamente da costituire ognuna un forte fattore selettivo per l'altra. In tal modo le due specie si adattano reciprocamente. Così, ad esempio, una modificazione intervenuta nella preda produce un corrispondente adattamento nel predatore e viceversa.
In alcuni casi il processo è così stretto che le specie diventano interdipendenti al punto che l'esistenza dell'una è univocamente condizionata dall'esistenza dell'altra e viceversa. Un esempio tipico è quello di alcune specie vegetali i cui fiori possono essere impollinati solo da una specie di insetti, i quali, a loro volta si cibano solo di quella specie vegetale. Nel caso in cui risultino associati organismi completamente dipendenti l'uno dall'altro si parla di "simbiosi obbligata" o "mutualismo".
La convergenza evolutiva si presenta quando specie diverse abitano lo stesso ambiente o ambienti simili. In tal caso le specie possono essere sottoposte a pressioni selettive dello stesso tipo e finiscono quindi per assomigliarsi, presentando un'analogia (non omologia) di forme e di strutture, senza derivare per questo da un antenato comune.
Un esempio classico di adattamento ad uno stesso ambiente, in questo caso il mare, si ha con lo sviluppo di una forma idrodinamica e di arti adatti at nuoto da parte di animali marini molto diversi come pesci, mammiferi (foche, balene, delfini), uccelli (pinguini) e rettili (ittiosauro).
In molti casi la convergenza evolutiva si verifica anche tra aree geografiche diverse, caratterizzate però da habitat e nicchie ecologiche sostanzialmente simili.
L'Australia offre senza dubbio l'esempio più eclatante di convergenza evolutiva. Questo continente, rimasto isolato dagli altri in tempi molto remoti, è stato colonizzato da mammiferi molto primitivi (monotremi e marsupiali), mentre negli altri continenti sono invece comparsi i più evoluti placentati.
I due gruppi hanno però occupato nicchie ecologiche quasi uguali ed oggi esistono degli analoghi in Australia dei principali mammiferi europei e americani. Molti di questi marsupiali hanno un aspetto così simile ai loro equivalenti placentati che i primi coloni europei li hanno chiamati con lo stesso nome, non essendo in grado di distinguerli.
Così il formichiere australiano presenta un muso allungato ed una lunga lingua vischiosa come il formichiere placentato ed il petauro volante è praticamente identico allo scoiattolo volante americano. Nonostante gli esempi siano molto numerosi esistono casi di divergenza. Ad esempio il canguro occupa in Australia la stessa nicchia ecologica occupata dagli equidi, senza per questo presentare con essi alcuna somiglianza.
Quando una struttura preesistente viene utilizzata per svolgere una funzione diversa da quella iniziale si dice che essa era preadattata alla nuova funzione ed il fenomeno prende il nome di preadattamento.
Certe strutture appaiono spesso troppo complesse per accettare l'idea che abbiano svolto fin dalla loro comparsa la funzione attuale. Per questo motivo si suppone che inizialmente svolgessero una funzione diversa e solo in un secondo momento si siano ulteriormente modificate per adattarsi ad una funzione diversa.
Il preadattamento è evidentemente un fatto casuale. Non possiamo certo distinguere all’interno degli esseri viventi odierni quali delle loro strutture attuali siano preadattate per svolgere un'altra funzione. Tuttavia alcune considerazioni possono essere fatte osservando le trasformazioni avvenute nelle linee filetiche dei fossili.
Così nell'evoluzione degli artropodi, prima che essi abbandonassero il mare, il primitivo rivestimento esterno di chitina aveva evidentemente una funzione di difesa contro i predatori. In seguito esso è stato utilizzato dagli artropodi terrestri per limitare l'evaporazione dell'acqua.
In modo analogo le branchie dei pesci avevano inizialmente solo una funzione trofica, filtrando l'acqua allo scopo di trattenere le particelle alimentari. Col procedere dell'evoluzione alcune pressioni selettive, legate probabilmente alla predazione, hanno determinato la comparsa di un tegumento protettivo più robusto, spesso provvisto di scaglie. Ciò però ha eliminato progressivamente la possibilità di rifornirsi di ossigeno direttamente dall'epidermide, creando una pressione selettiva in favore di altri meccanismi respiratori.
Il preadattamento viene considerato un importante meccanismo a disposizione della selezione naturale. D'altra parte è necessario rendersi conto che gli esseri viventi non sono progettati ex novo, partendo da zero, ma si evolvono a partire da strutture preesistenti.
Nei pesci primitivi ad esempio l'apertura boccale è fissa, non articolata in mascella e mandibola. In seguito le ossa che inizialmente si trovavano a proteggere le branchie vennero più convenientemente utilizzate per diventare mascelle articolate.
Nei rettili, discendenti dei pesci le mascelle sono formate da più ossa. Nei mammiferi, che discendono dai rettili, le mascelle sono formate da un unico pezzo. Nei fossili dei primi mammiferi si può osservare una progressiva miniaturizzazione delle altre ossa mascellari, che vengono infine utilizzate come ossicini nell'orecchio (staffa, incudine e martello).
Noi sentiamo con le ossa con cui i nostri antenati rettili masticavano e i rettili masticano con le ossa con cui i loro antenati pesci respiravano.
Ma non sempre le strutture preesistenti hanno modo di essere utilizzate. A volte certe strutture non trovano un'efficiente utilizzazione durante il processo evolutivo e spesso restano come caratteri vestigiali, a testimonianza del passato. Così l'uomo si ritrova con il coccige, termine con cui si indicano le ultime quattro vertebre saldate insieme, residuo della coda dei nostri antenati. L'appendice è un residuo di una sacca digestiva supplementare presente in alcuni erbivori, come il cavallo ed i roditori. Le ossa delle zampe posteriori della balena e quelle dei serpenti si trovano come piccoli ossicini all'interno del corpo dell'animale.
La piccola piega semicircolare che si trova nell'angolo interno dell'occhio umano è ciò che resta della terza palpebra, detta membrana nittitante, ancora ben sviluppata in molti anfibi, rettili ed uccelli, in cui svolge una funzione protettiva.
Modelli evolutivi: gradualismo filetico ed equilibri punteggiati
Il gradualismo filetico è il modello dominante all’interno della teoria sintetica. I suoi sostenitori affermano che i cambiamenti nella forma e nella struttura di un individuo si attuano lentamente ed impercettibilmente nel corso dell’evoluzione, come lo stesso Darwin aveva suggerito.
Ma se la trasformazione di una specie in un’altra fosse effettivamente un processo graduale e sfumato, i fossili dovrebbero documentarlo con numerose specie intermedie. In realtà, a tutt’oggi sono molto pochi i ritrovamenti fossili che documentano l’esistenza di linee evolutive in graduale trasformazione.
I biologi gradualisti attribuiscono la presenza di lacune (anelli mancanti) nelle linee filetiche a ‘buchi’ nella documentazione fossile. In altre parole gli anelli che ancora mancano per ricostruire gli alberi genealogici delle diverse specie, o non sono ancora stati scoperti, o non verranno mai scoperti perché non si sono verificate le condizioni adatte alla fossilizzazione.
Nel 1972 due paleontologi americani, Niles Eldredge e Stephen Jay Gould, ipotizzarono che la mancanza di forme intermedie non fosse da imputare alla incompletezza dei ritrovamenti, ma riflettesse effettivamente la vera natura del processo evolutivo.
La teoria degli equilibri punteggiati (o puntuati o intermittenti), da essi proposta, ipotizza che le specie rimangano stabili per lunghi periodi (periodi di equilibrio). La stabilità è però interrotta o “punteggiata” da modifiche rapide e radicali che portano infine alla comparsa improvvisa di una nuova specie.
Secondo i sostenitori di questo modello evolutivo, la maggior parte dei cambiamenti sarebbe concentrata nelle prime fasi della vita di una specie, in un lasso di tempo che equivale all’1% della durata della vita della specie in questione, secondo il modello di speciazione allopatrica che, come abbiamo visto, prevede una “rapida” speciazione nelle popolazioni isolate di piccole dimensioni.
È importante ricordare che, quando si parla di brevi periodi, ci si riferisce comunque a decine di migliaia di anni: la vita di una specie dura, infatti, mediamente alcuni milioni di anni.
Ad un’evoluzione lenta e graduale essi contrappongono quindi un’evoluzione a scatti.
Secondo questo nuovo modello le nuove specie si formerebbero in piccole popolazioni, accumulando rapidamente nuove modificazioni ed adattamenti, fino trasformarsi in una nuova specie.
A questo punto la nuova specie e quella vecchia rimarrebbero inalterate. Ma nel caso la nuova specie entri in competizione con la prima e risulti più adatta, dopo un breve periodo di coesistenza, la sostituisce decretandone l’estinzione.
In questo modo la teoria degli equilibri punteggiati sposta il processo selettivo a livello di specie. La selezione sarebbe in grado dunque di eliminare una specie perché meno adatta e di sostituirla con un’altra più adatta.
II processo di nascita selezione e morte dell’individuo, viene qui sostituito con quello di speciazione: selezione ed estinzione della specie.
Le prove dell’evoluzione
I biologi hanno accumulate un numero molto elevato di elementi che confermano la sostanziale validità della teoria evoluzionistica. Tra queste ricordiamo le più eclatanti, desunti da discipline diverse, a testimonianza del fatto che oramai la teoria sintetica dell’evoluzione rappresenta un comune punto di riferimento per gli studi biologici.
- paleontologia
Essendo oggi possibile procedere ad una datazione assoluta dei fossili, tramite rilievi radiometrici, si può facilmente constatare che i fossili più antichi appartengono ad esseri viventi meno complessi dei fossili più recenti. La complessità delle forme e delle strutture viventi è dunque una conquista evolutiva.
- anatomia comparata
Osservando organi omologhi di classi diverse di esseri viventi, è possibile, pur nell’apparente diversità. Rilevare una struttura comune, uno stesso progetto sviluppato con diverse variazioni sul tema, come se ciascuna classe si fosse sviluppata da un comune progenitore.
Le strutture omologhe sono parti di organismi di gruppi sistematici diversi che sono composte da elementi simili per forma e numero ed hanno rapporti anatomici simili. Se ad esempio analizziamo la struttura ossea interna degli arti dei vertebrati, scopriamo che il loro scheletro può essere ricondotto ad un unico modello formato da tre segmenti composti dagli stessi pezzi. I1 segmento prossimale (quello attaccato al corpo) è composto da un unico pezzo (omero/femore) il segmento mediano è composto di due pezzi (radio/tibia - ulna/perone); il segmento distale, che corrisponde alla mano e al piede è formato da un numero maggiore di ossa, variabile in funzione dei diversi adattamenti verificatisi.
Tutti i vertebrati hanno quattro arti e non sei, ne’ otto o venti. Tutti i mammiferi hanno sette vertebre cervicali. Se ogni animale fosse stato costruito ex novo non ci sarebbe ragione alcuna perché l’elefante debba presentare la stessa struttura corporea di un topo
- embriologia comparata
Confrontando embrioni differenti, ad esempio quelli delle classi dei vertebrati prima citati, nei primissimi stadi di sviluppo, si osserva una somiglianza impressionante. In tutti sono ad esempio presenti le fessure branchiali e la coda, anche in quelli che nella forma adulta non presentano tali caratteristiche.
Negli embrioni degli uccelli e delle balene, animali che da adulti sono privi di denti, compaiono degli abbozzi dentali che poi regrediscono.
Anche in tal caso si è indotti a pensare che il modello fondamentale della struttura corporea risalga ad un antico antenato comune.
Inoltre lo studio dell’embriologia comparata ha messo in evidenza come le strutture generiche, quelle comuni a molte specie e che permettono di stabilire l’appartenenza alle categorie sistematiche superiori (genere, famiglia etc.) compaiano prima di quelle specializzate, che determinano l’appartenenza ad una categoria inferiore.
Così nell’embrione umano compaiono prima le caratteristiche dei vertebrati, ad esempio l’abbozzo della colonna vertebrale, poi le caratteristiche dei mammiferi, come il pelo, ed infine le caratteristiche umane, come l’abbozzo del mento.
Tanto più lungo è il periodo dello sviluppo embrionale durante il quale embrioni di specie diverse si assomigliano, tanto più stretto si deduce essere il loro grado di parentela.
- citologia
Non può essere ritenuto un caso che, ad eccezione dei virus e dei procarioti, tutti gli esseri viventi siano costruiti tramite mattoni praticamente identici: le cellule. Le cellule di tutti gli eucarioti presentano infatti la stessa organizzazione strutturale e funzionale.
- biochimica
Anche le sostanze chimiche complesse, necessarie alla sopravvivenza degli esseri viventi sono praticamente le stesse per tutte le specie. Il codice genetico nel quale sono immagazzinate tutte le informazioni necessarie alla vita è universale: il significato delle triplette è lo stesso per tutti gli esseri viventi.
Le proteine risultano inoltre avere una struttura molto simile in gruppi sistematici molto vicini.
Evoluzione ed idea di progresso
I1 concetto di evoluzione ha sempre portato con sé l’idea di progresso, confermata in un certo qual modo dal fatto che gli esseri viventi che si sono succeduti nel tempo erano via via più grandi, più complessi e sofisticati.
Ciò è vero nella misura in cui l’evoluzione ha prodotto i risultati migliori rispetto a determinate condizioni ambientali, portando necessariamente modificazioni negli organismi al fine di un miglior adattamento al loro ambiente.
Ma è necessario tener presente che l’evoluzione è un processo casuale, che agisce attraverso la selezione naturale su materiale già presente e non sempre ottimale per essere adattato alle diverse e nuove esigenze ambientali.
Così, oltre ad organi perfettamente adattati che vengono proposti come esempi della perfezione raggiunta dall’evoluzione (l’occhio ad esempio), vi sono strutture poco efficienti che probabilmente un ingegnere o un architetto ridisegnerebbero completamente.
Molti problemi umani come le difficoltà a partorire o la tendenza a soffrire di mal di schiena possono essere fatte risalire, ad esempio, al passaggio alla stazione eretta di una struttura scheletrica precedentemente adattatasi alla postura quadrupede.
Gould, il teorico degli equilibri intermittenti, afferma che la presenza di queste ‘imperfezioni’ tra gli esseri viventi sono prove molto più convincenti dell’evoluzione di quanto lo siano gli esempi solitamente citati di adattamenti perfetti.
Inoltre, visto che l’evoluzione tende a modificare una specie in modo da raggiungere il miglior compromesso possibile tra esigenze ambientali ed una serie di limitazioni strutturali ereditate dalla storia evolutiva passata dell’organismo, dobbiamo affermare che tutte le specie, per il fatto di esistere, sono attualmente il miglior esempio di adattamento alla propria nicchia.
Dunque poiché tutte le specie sono adattate alla loro nicchia è difficile in biologia dire qual è l’essere vivente più progredito.
E’ più progredita una scimmia o l’uomo? Se pensiamo allo sviluppo del cervello, certamente l’uomo, ma se pensiamo alla capacità di sopravvivere in un foresta, certamente la scimmia.
In definitiva in biologia l’idea di progresso è un concetto di difficile applicazione, che può facilmente essere frainteso o dar luogo a false interpretazioni.
Per questo motivo al termine progresso si sostituiscono termini più tecnici e di significato più limitato o convenzionalmente definito, come i termini “semplice/complesso” o “primitivo/evoluto”.
Un organismo si dice più complesso di un altro quando, in riferimento alla loro organizzazione strutturale e funzionale , sono rilevabili un maggior numero di parti differenziate.
I termini “primitivo/evoluto” descrivono il livello di trasformazioni che un organismo o una sua parte hanno subito rispetto all’antenato. Così un batterio è un organismo primitivo essendo rimasto praticamente immutato da quando è comparso sino ad oggi. D’altra parte la zampa di un cavallo è più evoluta rispetto alla mano dell’uomo, rimasta praticamente immutata nel corso dell’evoluzione. La zampa del cavallo ha infatti subito un maggior numero di modificazioni e si è maggiormente specializzata a compiere una funzione. Esistono esseri viventi che, essendosi adattati ad ambienti che non hanno subito praticamente alcuna modificazione da milioni di anni, sono rimasti immutati per periodi altrettanto lunghi.
Si tratta di specie primitive a volte chiamate fossili viventi, come il limulo, un artropode molto simile agli antichi trilobiti ora estinti, o la Ginkgo biloba, unagimnosperma rimasta praticamente uguale da 200 milioni di anni.
Fonte: http://rexmagister.altervista.org/downloads/BiologiaDOC.zip
sito web: http://rodomontano.altervista.org/biologia.php
Autore del testo: Rodomontano
Biologia appunti parte 4
Visita la nostra pagina principale
Biologia appunti parte 4
Termini d' uso e privacy