Cellule animali e cellule vegetali
Cellule animali e cellule vegetali
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
LE STRUTTURE E LE FUNZIONI DELLE CELLULE
DEFINIZIONE E CENNI STORICI
Tutti gli esseri viventi sono costituiti da unità elementari: cellule. Ci sono alcuni, i più semplici, sono formati da una sola cellula (organismi unicellulari); altri sono molto più complessi e hanno moltissime cellule (organismi pluricellulari). Questi ultimi posseggono cellule che non sono tutte uguali tra loro, ma ci sono alcune specifiche differenze nella forma, nelle dimensioni, nelle funzioni particolari. All'interno del corpo di un organismo pluricellulare i sistemi di cellule differenziate (simili tra loro per origine embrionale, per forma e per funzione) e tutti i materiali che elaborano sono chiamati tessuti, questi unendosi opportunamente fra loro danno origine a strutture con un livello di organizzazione funzionale più complesso chiamate organi.
Lo studio dei tessuti fu chiamato da Meyer istologia( dal greco histos = trama, tessuto e logos =discorso). Il termine utilizzato in biologia non è propriamente adatto: a parte qualche tessuto animale, come il connettivo fibroso, raramente l'organizzazione dei tessuti biologici ricorda la struttura della stoffa. Siccome il termine ha avuto un certo successo lo si usa ancora. L'istologia si propone di studiare la struttura, l'ultrastruttura dei tessuti e il loro significato funzionale e i meccanismi che sono alla base della loro funzione. Un'altra branca della biologia che non può assolutamente mancare è la citologia (dal greco kylos =cella, cavità e logos =discorso) che partendo agli inizi a descrivere solo la morfologia, ora il suo campo d'indagine e sperimentazione si è esteso alle funzioni delle cellule e ai suoi meccanismi, fino ad esplorare l'organizzazione molecolare delle sostanze organiche.
Lo sviluppo di questi due settori d'indagine biologica è legata alla storia del microscopio: strumento che consente l'osservazione di strutture enormemente piccole.
Strumenti per ingrandire gli oggetti (lenti o bocce di vetro piene d'acqua) venivano già utilizzati nel medioevo, specie tra i mercanti di stoffe e i tagliatori di diamanti. Ma fu solo a partire dal XVIII secolo che, prima la lente (semplice microscopio) e poi il microscopio composto, vennero impiegati in maniera sistematica per osservare gli organismi viventi.
Nel 1665 il fisico inglese Robert Hooke (1653-1702) introdusse il termine “cellula”, esaminando al microscopio alcune sezioni di sughero. Osservò che il sughero aveva un aspetto simile a un favo di api, formato da un insieme di piccolissime cavità regolari circondate da una parete a cui diede il nome di cells (cellette). Egli aveva osservato solo l'involucro, la parte cellulare di elementi morti, svuotati del loro contenuto. Dopo qualche decennio Antoni van Leeuwenhoek, naturalista olandese (1632-1723) e fondatore dell'istologia, fece una importante scoperta nell'acqua stagnante: vide la presenza di alcuni piccoli animaletti (animalcula), chiamati in seguito infusori. Quindi si scoprirono le prime cellule animali. Per tutto il XVIII secolo si moltiplicarono molte osservazioni, ma nessuno aveva ancora compreso il vero significato di queste strutture. Stava crescendo tuttavia l'ipotesi che l'organizzazione degli animali era in un certo senso simile a quella dei vegetali. Si spostò sempre più l'attenzione dalla parete a quello che vi era contenuto all'interno di queste strutture (cellule). Nel 1780 lo studioso italiano Fontana, mentre osservava cellule epiteliali dell'anguilla, scoprì al loro interno delle strutture più scure chiamate nucleo. La presenza di un organello comune a tutte le cellule vegetali venne scoperta nel 1831 dal naturalista scozzese Robert Brown (1773-1858).
Il primo ad avanzare e sostenere l'ipotesi che tutti gli organismi viventi nascono da cellule e sono formati da cellule o vescicole è stato il tedesco Oken nel 1805 (1ª teoria cellulare). Nel 1838 fu la volta del botanico Schleinden ad affermare che tutti gli organismi vegetali sono costituiti da cellule. L' anno successivo (1839) lo zoologo Schwann affermò lo stesso per gli organismi animali, usando la prima volta l'espressione teoria cellulare con la nascita della citologia (disciplina che studia la morfologia delle cellule), stabilendo che:
- la cellula è l'unità funzionale degli organismi animali e vegetali
- esiste una relazione tra le differenti forme delle cellule e le loro funzioni,
- riconosciuta una organizzazione, cioè le cellule non erano giustapposte ma nell'organismo collaborano in un sistema unitario,
- le cellule sono capaci di assorbire dall'ambiente selettivamente le sostanze di cui necessitano
Con l'apporto di tanti altri contributi dovuti da numerosi scienziati che hanno operato in tutta la seconda metà dell'Ottocento, si perfezionò e si diffuse la teoria cellulare, riconosciuta in maniera definitiva verso la fine del'800:
- tutte le parti degli organismi vegetali e animali sono costitute da cellule o da materiali da esse prodotti
- le cellule non si formano ex novo, ma derivano da altre cellule (Rudolf Virchow 1821-1902: omnis cellula e cellula
- gli organismi pluricellulari che hanno riproduzione sessuale derivano dall'uovo fecondato (zigote) che è una cellula
- la cellula, unità fondamentale, strutturale e funzionale, possiede in sé tutti gli attributi e le capacità che caratterizzano i viventi e che li distinguono da ciò che non è vivente
L'affermazione della teoria ebbe ripercussioni importanti in tutti i campi della biologia. Nello stesso periodo inoltre attraverso l'osservazione al microscopio sono stati individuati e descritti tutti i principali organuli cellulari visibili al microscopio ottico.
Cellule vegetali
Forme, dimensione e struttura delle cellule vegetali sono variabili in base alle funzioni che esse svolgono all'interno dell'organismo. Le leggi della natura limitano le dimensioni cellulari. Ci sono cellule molto piccole il cui diametro è intorno a 0,5 μm (alcune alghe azzurre) e cellule molto più grandi appartenenti ad alcune alghe verdi del genere Valonia che hanno diametri di diversi centimetri (sono atipiche perché nel citoplasma si trovano contemporaneamente più nuclei – stato cenocitico cioè comunità cellulare). La maggior parte delle cellule vegetali delle piante superiori ha dimensioni più modeste e i loro diametri sono tra i 20 e i 350 μm. Alcune cellule di lino, come le fibre sclerenchimatiche (tessuto meccanico di piante superiori), raggiungono lunghezze di circa 10 cm: presentano pareti ispessite. A determinare le dimensioni della cellula ha particolare importanza il rapporto superficie/volume, da cui dipende l'efficacia degli scambi fra la cellula e l'ambiente che lo  circonda. Le leggi di natura limitano le dimensioni cellulari.
circonda. Le leggi di natura limitano le dimensioni cellulari.
Le cellule non possono avere dimensioni né troppo grandi né troppo piccole: devono essere abbastanza grandi da contenere una certa quantità di DNA, di molecole proteiche e di strutture interne necessarie alla loro sopravvivenza e riproduzione, le dimensioni nel contempo sono limitate dalla necessità di avere un'area superficiale abbastanza estesa da poter introdurre un'adeguata quantità di sostanze nutritive dall'ambiente esterno ed eliminare le sostanze di rifiuto. Le cellule più grosse hanno un'area superficiale maggiore rispetto a quelle piccole, ma un rapporto superficie/volume minore.
Esempio: un'unica grande cellula che ha per ipotesi una forma cubica con un lato pari a 30 μm avrà un volume complessivo pari a volume = 30 μm x 30 μm x 30 μm = 27.000 μm3
Se nello stesso volume poniamo di avere un certo numero di cellule cubiche con lato pari a 10 μm avremo appunto sempre lo stesso volume complessivo, ma le aree superficiali complessive sono molto differenti:
- area del cubo grande = 6 x(30 μm x 30 μm) 5.400 μm2
- area del cubo piccolo = 6 x(10 μm x 10 μm) 600 μm2
poiché i cubi piccoli sono 27, l'area superficiale totale è = 27 x 600 μm2 = 16.200 μm2
Se il volume citoplasmtico aumenta in maniera eccessiva il nucleo all'interno della cellula non riuscirà più a controllare il metabolismo cellulare e gli scambi con l'ambiente risulteranno inadeguati per mantenere il livello metabolico ottimale.
 Riguardo alla forma occorre dire che essa varia in funzione delle funzioni e del numero di cellule che costituiscono l'organismo. Gli individui unicellulari hanno cellule per lo più sferiche; mentre negli organismi pluricellulari le cellule hanno aspetti tipici con strutture rigide, più o meno poliedrici e allungati, che sono determinati durante la fase di differenziazione in relazione alla funzione che svolge nell'organismo. Durante il differenziamento si assiste alla formazione di un certo numero di tipi di cellule, a partire da tessuti meristematici, privi di qualsiasi specializzazione.
Riguardo alla forma occorre dire che essa varia in funzione delle funzioni e del numero di cellule che costituiscono l'organismo. Gli individui unicellulari hanno cellule per lo più sferiche; mentre negli organismi pluricellulari le cellule hanno aspetti tipici con strutture rigide, più o meno poliedrici e allungati, che sono determinati durante la fase di differenziazione in relazione alla funzione che svolge nell'organismo. Durante il differenziamento si assiste alla formazione di un certo numero di tipi di cellule, a partire da tessuti meristematici, privi di qualsiasi specializzazione.
Tutte le cellule sono separate dall'ambiente esterno per mezzo di una membrana detta plasmalemma, semipermeabile e capace di una certa selezione e controllo del movimento delle sostanze. I vegetali oltre al plasmalemma più esternamente posseggono una parete che ne costituisce il supporto meccanico.
Cellule animali
Come nelle cellule vegetali anche le cellule animali presentano dimensioni che possono variare in range molto ampi: di norma non sono visibili ad occhio nudo, se si eccettuano alcuni protisti e le cellule uovo di alcuni animali. Le cellule più piccole sono alcune degli uccelli con diametro pari 2 μm e sempre fra gli uccelli si trovano cellule fra le più grandi con diametro di 85 mm come la cellula uovo di struzzo. Le cellule hanno dimensioni piuttosto costanti per ogni tipo: tra i vertebrati ci sono specie a cellule piuttosto grandi come quelle degli anfibi, oppure molto piccole come gli uccelli, i pesci e i mammiferi. In animali appartenenti alla stessa specie, ma di mole molto diversa, non cambiano le 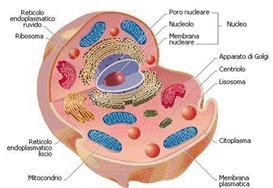 dimensioni delle cellule, ma il loro numero: legge della costanza del volume cellulare
dimensioni delle cellule, ma il loro numero: legge della costanza del volume cellulare
Anche per le cellule animali valgono le stesse considerazioni sul rapporto superficie/volume.
Riguardo alla forma possiamo osservare che anche qui vale il discorso se l'organismo è unicellulare o pluricellulare. Negli organismi con una sola cellula, questa unica cellula ha la forma dell'individuo ed è caratteristica della specie. Questa cellula può avere una forma fissa e complicata dalla presenza di appendici di vario genere (ciglia, flagelli) o variabile come nelle amebe. Negli organismi pluricellulari la forma delle cellule non ha rapporti con la forma dell'individuo, ma è direttamente legata alla funzione, al grado di specializzazione raggiunto e alla eventuale azione meccanica esercitata dalle altre cellule. C'è sempre uno stretto legame fra forma e funzione, esempi:
- cellule muscolari, specializzate nella contrazione hanno forma allungata e fusiforme
- cellule nervose che devono prendere rapporto con altre cellule anche a distanza per ricevere lo stimolo e trasmettere una risposta, presentano prolungamenti diversi anche molto estesi
Membrana plasmatica (plasmalemma)
Prima di iniziare il nostro viaggio all'interno della cellula è opportuno descrivere il suo costituente più periferico, cioè la membrana citoplasmatica che rappresenta il confine tra l'ambiente interno e quello esterno.
La membrana plasmatica è un sottile involucro di circa 7-8 nm (1 nm = 10-9) di spessore,
 che avvolge la cellula separando completamente il citoplasma dall'ambiente esterno. È visibile solo al microscopio elettronico, mentre al microscopio ottico è possibile riconoscere solo i limiti della cellula.
che avvolge la cellula separando completamente il citoplasma dall'ambiente esterno. È visibile solo al microscopio elettronico, mentre al microscopio ottico è possibile riconoscere solo i limiti della cellula.
 La membrana plasmatica si interpone tra le possibili modificazioni dell'ambiente esterno e il citoplasma, ma non ne costituisce solo il confine fisico della cellula, essa svolge anche molteplici funzioni.
La membrana plasmatica si interpone tra le possibili modificazioni dell'ambiente esterno e il citoplasma, ma non ne costituisce solo il confine fisico della cellula, essa svolge anche molteplici funzioni.
Tra le più importanti:
- la permeabilità selettiva, che permette di regolare gli scambi tra cellula e ambiente
- coordinamento delle attività metabolica della cellula per la presenza di numeri enzimi
- agisce come superficie sensibile per la presenza di numerose molecole recettrici in grado di rispondere a stimoli specifici o a stimoli di carattere generale
La membrana non è una struttura rigida e fissa, ma dinamica, perché le sue componenti molecolari vengono continuamente rinnovate e possono modificare la loro disposizione in relazione alle funzioni che la membrana svolge. Può variare anche la sua estensione.
La membrana è asimmetrica, perché presenta componenti diverse e variamente distribuite su ciascuna delle sue due facce, extra e intracellulare, presentando proprietà funzionali diverse su entrambe le facce.
I componenti chimici sono lipidi, proteine e carboidrati che interagiscono fra loro in maniera coordinata. I lipidi costituiscono il 40% in peso della membrana: rappresentati fondamentalmente da lipidi complessi tra cui predominano i fosfolipidi, poi colesterolo, glicolipidi e talvolta piccole quantità di trigliceridi. I fosfolipidi sono molecole a doppia polarità (anfipatiche) con un polo (testa) idrofilo e un polo (coda) idrofobo.
Proprio grazie a questa caratteristica i lipidi si organizzano nelle membrane biologiche con un orientamento preciso formando un doppio strato di molecole di circa 5 nm di spessore, nel quale le estremità idrofobe di uno strato di molecole affiancate si affacciano a alle estremità idrofobe delle molecole di uno strato simile. Mentre le teste idrofile sono rivolte rispettivamente verso l'ambiente extracellulare o verso il citoplasma.
Inoltre questo doppio strato si comporta come un sistema relativamente fluido nel quale le molecole possono muoversi spostandosi lateralmente.
Le proteine (circa il 60% in peso) possono essere semplici o coniugate e di peso molecolare variabile. Alcuni tipi di proteine sono presenti nelle membrane di tutte le cellule, mentre altri sono specifici per tipi di cellule. Le proteine svolgono diverse funzioni:
- funzione enzimatica con ruolo nei fenomeni di trasporto
- recettori
- anticorpi
La disposizione non è omogenea e interagiscono più o meno fortemente con i lipidi
I carboidrati (intorno al 2% del peso) sono polimeri con breve catena (oligosaccaridi) a composizione molto eterogenea e si legano per lo più alle proteine nel versante extracellulare e qualche volta alla testa idrofila dei fosfolipidi.
Il modello di organizzazione molecolare della membrana che consente di interpretare le diverse caratteristiche morfologiche e funzionali è stato proposto nel1972 da Singer e Nicholson. Il modello rappresenta un mosaico fluido in cui uno strato bimolecolare di lipidi forma l'impalcatura alla quale le proteine aderiscono in superficie (proteine periferiche o estrinseche), trattenute da forze di attrazione polare, o sono inserite tra i lipidi (proteina integrale o intrinseca) e attraversano tutto o in parte il doppio strato lipidico, trattenute da forze non polari. Avendo il doppio strato lipidico un certo grado di fluidità le molecole proteiche e quelle lipidiche possono diffondersi nel piano della membrana e talvolta spostarsi nel suo spessore.
PROCARIOTI ED EUCARIOTI
L'atmosfera primordiale e le prime cellule
La vita è un processo materio-energetico iniziato circa 3,5miliardi di anni fa. Le prime reazioni chimiche si basavano sull'idrogeno e pochi altri elementi (ossigeno, carbonio, azoto..). Molto probabilmente l'atmosfera della Terra qualche miliardo di anni fa era simile a a quelle di alcuni pianeti gioviani: assenza di ossigeno, presenza di vapor acqueo, e ricchezza di idrogeno, azoto, ammoniaca, anidride carbonica e metano. Questi elementi e molecole primordiali in presenza di alte energie e scariche elettriche dei fulmini e di abbondanti radiazioni ultraviolette (assenza del strato di ozono) si aggregarono in molecole più grosse che a loro volta reagirono tra loro per formare le prime molecole organiche. Questa ipotesi fu formulata la prima volta da Oparin- Holdane negli anni 40: essi affermarono che le molecole organiche poste alla base della vita sulla Terra (acidi nucleici, proteine, carboidrati...) si sono formate per aggregazioni successive a partire dalle prime sostanze presenti nell'atmosfera. Stanley Miller diede prova e sostegno a questa ipotesi con l'esperimento del 1953: mise in un recipiente vapor acqueo, idrogeno, metano e ammoniaca creando condizioni di alta pressione e sottoponendo la miscela a scariche elettriche, cercando di ricreare le possibili condizioni della atmosfera primordiale. Ebbe un risultato sorprendente: ottenne la sintesi di vari amminoacidi.
L'enorme varietà di forme di organismi viventi, dai più piccoli ai più grandi, presenta una base comune: come abbiamo già visto, tutti gli esseri viventi sono costituiti da cellule. Ci sono organismi che contengono una sola cellula, come i Batteri e i Protozoi: altri che invece ne contengono moltissime come gli organismi pluricellulari come l'uomo.
Sulla base dell'analisi di alcuni reperti fossili risulta che le prime cellule apparvero sul nostro pianeta circa 3 miliardi di anni fa. Queste cellule erano prive di nucleo.
Prima della comparsa delle prime cellule certamente c'è stato un lungo periodo in cui comparvero le prime molecole di carbonio e i precursori degli amminoacidi, le prime basi azotate e i carboidrati. Quindi per polimerizzazione si formarono le macromolecole. Formazione delle prime proteine e quindi anche dei primi enzimi. Successivamente le molecole si organizzarono in coacervati: parte interna fluida e una superficie esterna molto simile, nella composizione chimica, a quella della membrana cellulare.
I primi procarioti: cellule piccole e struttura molto semplice
Si formarono quindi le prime macromolecole costituite da acidi nucleici determinando la comparsa del corredo genetico che permise la prima forma di vita capace di autoriprodursi. Le prime cellule quindi avevano un'organizzazione cellulare molto semplice, perché erano prive di un vero e proprio nucleo. Nei procarioti le molecole di DNA e RNA sono mescolate con altre sostanze presenti nel citoplasma. I primi procarioti probabilmente impiegavano quale fonte di energia altre molecole organiche, perciò erano eterotrofi e anaerobi, perché l'atmosfera era priva di ossigeno. Nel processo evolutivo ad un certo punto subentra la capacità di sintetizzare altre macromolecole, pigmenti fotosintesi e trasformò i primi procarioti eterotrofi in organismi autotrofi, cioè capaci di produrre, attraverso la fotosintesi, gli zuccheri necessari per il nutrimento. Il processo fotosintetico determinò quindi la presenza di ossigeno nell'atmosfera. e portò alla presenza di procarioti autotrofi aerobi.
Si fa risalire a circa 1 miliardo di anni fa la comparsa sulla Terra di cellule dotate di un nucleo vero e proprio circondato da membrana, cioè organismi eucarioti
I procarioti hanno n a struttura cellulare molto più semplice rispetto a quella degli eucarioti. I procarioti sono privi di un'apprezzabile compartimentazione funzionale dello spazio intracellulare: sono costituiti da una membrana cellulare ( avvolte protetta da parete cellulare esterna), contenente citoplasma con acidi nucleici, proteine, ecc.
I procarioti si dividono in Archeobacteria che vivono in condizioni estreme, nelle profondità abissali degli oceani, nelle vicinanze delle bocche idrotermali e vulcaniche, laghi salati, dove i fattori abiotici (pressione, temperatura, pH, salinità,...) hanno valori estremi rispetto a quelli di ambienti dove vivono la maggior parte degli organismi viventi. Ci sono tra gruppi principali di Archeobacteria in base alla sostanza impiegata per ricavare energia:
1. metanobatteri che convertono idrogeno e anidride carbonica in metano
2. sulfobatteri che estraggono energia dai composti dello zolfo
3. alobatteri che vivono in ambienti fortemente salini
Molto verosimile che l'evoluzione biologica abbia avuto inizio a partire da organismi di questo tipo, per il semplice motivo che miliardi di anni fa le condizioni ambientali del pianeta in termini di temperature, pressione ecc.. dovevano essere simili a quelle riscontrabili negli habitat dove vivono gli archeobatteri.
Gli Eubacteria comprendono moltissime specie di batteri conosciuti, da quelli fotosintetizzanti (alghe azzurre) a quelli patogeni.
La forma della cellula procariote costituisce un utile criterio per la classificazione: essa è almeno di quattro tipi: bacilli, cocchi, vibrioni e spirali.
Nel batterio tipo il citoplasma non è omogeneo, ma presenta alcuni granuli costituiti da materiale di accumulo (sostanze energetiche). Il citoplasma rappresenta per l'organismo procariote una sorta di fattoria chimica, dove avvengono tutte le reazioni chimiche (metabolismo): alcune si svolgono impiegando sostanze organiche già presenti per ricavare energia (catabolismo); altre portano alla formazione di nuove sostanze a partire da molecole più semplici preesistenti (anabolismo). Il DNA è mescolato all'interno del citoplasma ed è libero senza confinamento funzionale in un nucleo. La riproduzione cellulare avviene per scissione, dopo che il DNA si è duplicato.
Se ci sono condizioni ambientali ottimali ( risorse energetiche e optimum di temperatura e altri fattori abiotici) i procarioti si riproducono molto velocemente (ogni 10-20 minuti): in poco meno di 12 ore da un batterio se ne possono avere ben 4 miliardi. Quando le condizioni sono sfavorevoli (assenza di risorse nutritive e fattori abiotici non ottimali) i batteri possono formare delle cellule molto resistenti dette spore che gli consentono di superare il momento sfavorevole: alcune spore batteriche sono in grado di resistere anche a temperature di ebollizione dell'acqua.
I procarioti vivono in diversissimi ambienti: nei suoli, nelle acque, nell'aria e sulle superfici di qualsivoglia vivente più grande di loro. Moltissimi batteri sono patogeni dell'uomo. Altri batteri invece sono utili all'uomo con cui stabiliscono relazioni simbiontiche, come alcune specie che sono ospitate dall'intestino e che sintetizzano la vitamina K.
Eucarioti: cellule divise in compartimenti che svolgono funzioni diverse
Le cellule di molte alghe, dei Muschi, delle Felci, delle piante e degli animali esaminate al microscopio ottico appaiono trasparenti per la quasi totalità del corpo cellulare, tranne che per una piccola parte scura centrale che rappresenta il nucleo che è una struttura altamente rifrattiva. Tutte le cellule eucariotiche hanno questa struttura avvolta da una doppia membrana, al cui interno vi è il DNA. Come si è visto precedentemente, queste cellule possono avere forme e dimensioni differenti, in relazione alla loro differenziazione funzionale, ma tutte presentano un nucleo ben individuabile e diversi altri organelli nel citoplasma. Ogni organello, come vedremo meglio più avanti, svolge una funzione particolare in una complessa architettura fatta a compartimenti. Fra tutti gli organuli ci sono alcuni come i mitocondri, nella cellula animale e vegetale, e i cloroplasti nella sola cellula vegetale, che posseggono un proprio DNA, pertanto capaci di autoriprodursi. La cellula procariote e quella eucariote si differenziano per l'assenza o presenza di nucleo, mitocondri, cloroplasti e di altri organuli e in particolare di microtubuli e microfilamenti che contribuiscono a definire un citoscheletro, che aiuta alla tenuta della struttura generale della cellula e le conferisce una certa elasticità.
Procarioti Eucarioti
Organismi Batteri e cianobatteri Protisti, Funghi, Piante e Animali
Dimensioni cellulari Generalmente da 1 a 10 m di dimensioni lineari Generalmente da 5 a 100 m di dimensioni lineari
metabolismo Anaerobio o aerobio aerobio
organelli Pochi o nessuno Nucleo, mitocondri, cloroplasti, reticolo, endoplasmatico, ecc..
DNA DNA circolare nel citoplasma Molecole molto lunghe di DNA lineare contenenti molte regioni non codificanti; circondato da un involucro nucleare
RNA e proteine RNA e proteine sintetizzate nello stesso compartimento RNA sintetizzato ed elaborato nel nucleo; proteine sintetizzate nel citoplasma
Citoplasma Assenza di citoscheletro: niente flussi citoplasmatici, endocitosi ed esocitosi Citoscheletro composto da filamenti proteici; flussi citoplasmatici; endocitosi e esocitosi
Divisione cellulare Cromosomi separati mediante attacco alla membrana plasmatici Cromosomi separati da un fuso di citoscheletro
Organizzazione cellulare In genere unicellulare In genere multicellulare, con differenziamento di molti tipi cellulari
Nella cellula procariote la quasi totalità delle funzioni cellulari avviene nel citoplasma e nella membrana cellulare. Nella cellula eucariote sono presenti unità strutturali distinte, dette organuli, separati dal resto della cellula da sistemi più o meno complessi di membrane che sono in grado di determinare una compartimentazione delle varie funzioni cellulari, quindi fisicamente separate. Nel citoplasma c'è una rete di membrane che risultano tra loro anastomizzate e derivanti da invaginazioni della membrana esterna dell'involucro nucleare (reticolo endoplasmatico). Altro aspetto esclusivo della cellula eucariote è la capacità di scambiare materiali tra i diversi compartimenti della cellula e l'esterno della stessa: capacità della membrana cellulare di espellere attraverso l'esocitosi sostanze di rifiuto e di assumere attraverso l'endocitosi le sostanze utili alle funzioni cellulari. L'esocitosi è possibile perché ci sono porzioni di membrana che avvolgono sostanze presenti nella cellula (esempio proteine contenute in vescicole) che si fondono con la membrana cellulare, rilasciando all'esterno della cellula il loro contenuto. Mentre l'endocitosi avviene perché parti di membrana cellulare si introflettono (invaginandosi) avvolgendosi su se stesse formando delle vescicole contenenti sostanze che in precedenza si trovavano all'esterno.
Il citoplasma
Compartimento citoplasmatico: citoplasma all'interno della membrana plasmatica; parte prevalente della massa cellulare. Dal punto di vista chimico: acqua con circa il 75%; proteine con 15-20%; altri composti organici (zuccheri, acidi grassi, nucleotidi, .....) 5-10%. Il citoplasma al microscopio ottico, a forte ingrandimento, veniva considerato come un sistema colloidale privo di organizzazione strutturale, assumendo aspetto granulare, reticolare o vacuolare a seconda del sistema di fissazione e preparazione del vetrino. Si è dovuto attendere l'uso del microscopio elettronico per rilevare un'organizzazione strutturale molto complessa all'interno del citoplasma.
La struttura dell'interno cellulare è costituita per lo più da un intricato sistema di membrane che forma un labirinto tridimensionale di cavità, in cui si distinguono diverse sezioni collegate fra loro e con l'involucro nucleare. Le membrane interne in pratica ripropongono la stessa organizzazione molecolare della membrana plasmatica, con la presenza di un mosaico fluido determinato dalle stesse categorie di molecole organiche (fosfolipidi e proteine).
Nel citoplasma si distinguono:
citosol matrice citoplasmatica o citoplasma fondamentale esterna alle membrane. Costituisce il vero ambiente interno delle cellule. Nella matrice sono presenti una serie di elementi citoscheletrici rappresentati da microtubuli, microfilamenti e microtrabecole che costituiscono un'impalcatura scheletrica dinamica che favorisce il mantenimento delle forma, partecipa ai fenomeni di movimento e fa da supporto alle migliaia di proteine solubili, agli enzimi e ai ribosomi, consentendo in tal modo lo svolgimento delle razioni enzimatiche, cioè alle attività di sintesi e di degradazione di molecole (metabolismo intermedio). I microtubuli sono strutture tubulari abbastanza rigide con pareti formate da unità globulari di una proteina detta tubulina. Le diverse aggregazioni dei microtubuli entrano in gioco in importanti processi cellulari, ad esempio nella separazione dei cromosomi durante la divisione nucleare. Sono anche alla base delle appendici come le ciglia e i flagelli. I microfilamenti più sottili sono formati da una proteina detta actina che associata ad altre proteine, come la miosina, permettono la contrazione, quindi i movimenti citoplasmatici e dell'intera cellula.
Con la microscopia elettronica, come si è già detto, si è individuato il sistema di endomembrane costituito da tre sezioni molto interconnesse fra loro, anche se morfologicamente distinte: reticolo endoplasmatico nella forma granulare (o rugoso), reticolo endoplasmatico nella forma liscia e l'apparato del Golgi
Ora inizieremo ad esaminare una per una tutte le strutture interne di una cellula eucariote.
Reticolo endoplasmatico
La porzione più sviluppata del sistema di membrane. Abbiamo già visto che esistono due forme: queste sono presenti in tutte le cellule e la loro espansione varia dal tipo di cellula e dallo stadio di vita della cellula stessa. Il reticolo funziona come una specie di sistema circolatorio per la distribuzione intracellulare di sostanze diverse, in quanto le sue membrane sono dinamiche e possono assumere stati di continuità funzionale con la membrana plasmatica, con l'involucro nucleare e con l'apparato del Golgi, attraverso questa serie di connessioni è possibile il trasporto di particelle e ioni verso l'esterno e dall'esterno della cellula o lo scambio di materiali con il nucleo. Reticolo endoplasmatico rugoso dà origine a membrane e proteine: per la presenza di piccole strutture rotondeggianti, chiamate ribosomi, che aderiscono alla faccia citoplasmatica delle membrane. I ribosomi sono presenti nel citoplasma in forma libera, come nei procarioti, oppure liberi e legati al reticolo endoplasmatico negli eucarioti. Hanno un'importante funzione nella sintesi proteica, in quanto costituiscono il supporto fisico durante l'assemblaggio. I ribosomi sono gli unici organelli che non sono circondati da membrana e sono molto piccoli e distinti in due subunità: maggiore e minore entrambe costituite da un'associazione fra RNA e proteine. Il reticolo endoplasmatico rugoso forma un sistema di cavità per lo più appiattite (sacche o cisterne), spesso molto ordinate, qualche volta con lume virtuale e addensate in alcune zone della cellula. La funzione più importante si ricollega alla sintesi delle proteine che devono essere secrete, oppure segregate all'interno delle cellule nei lisosomi (vedasi in seguito). Le proteine vengono sintetizzate a livello dei ribosomi aderenti alle membrane e passano nelle sue cavità e vengono quindi da qui convogliate verso l'apparato del Golgi (vedasi in seguito) e poi all'esterno. Reticolo endoplasmatico liscio non presenta ribosomi, forma un sistema di cavità tubulari molto intrecciate fra loro e apparentemente disordinato. Forma delle connessioni con il reticolo endoplasmatico rugoso; partecipa alla demolizione di sostanze inutili o nocive o di farmaci (detossificazione), al metabolismo del glicogeno e al metabolismo dei lipidi.
Apparato del Golgi rielabora, seleziona e trasporta i prodotti cellulari
Attraverso la microscopia elettronica è stato possibile individuare la presenza dell'apparato in tutte le cellule eucariotiche, dopo che fu scoperto per la prima volta da Golgi Camillo (1843-1926) nelle cellule nervose. La sua struttura vede la presenza di sacchetti appiattiti (cisterne) disposti in pile cui si associano vescicole (piccole bolle provenienti dal distacco di porzioni marginali delle cisterne stesse). Le pile di cisterne possono essere numerose e disposte in prossimità del nucleo. La funzione dell'apparato del Golgi consiste in una sorte di stazione di elaborazione, sintesi e smistamento di materiali in connessione con il reticolo endoplasmatico e la membrana plasmatica. Il complesso permette un continuo flusso di materiali, che possono essere stati sintetizzati in altri distretti della cellula e qui vengono rielaborati e modificati, passando nelle vescicole che si distaccano dai margini delle cisterne, questi materiali giungono fino alla membrana plasmatica e quindi vengono secrete.
Lisosomi demoliscono le sostanze alimentari e di rifiuto delle cellule
Organuli delimitati da una membrana e contengono moltissimi enzimi idrolitici. Sono caratterizzati da un accentuato polimorfismo e svolgono importanti funzioni di demolizione grazie alla presenza di enzimi idrolasi. Possono agire su quasi tutte le categorie di sostanze biologiche. Digestione intracellulare di materiali assunti dall'esterno il lisosoma si fonde con una vescicola di fagocitosi o di pinocitosi formando un vacuolo digestivo nel quale il materiale ingerito viene a contatto con gli enzimi e demolito in molecole a basso peso molecolare che vengono assorbite attraverso la membrana del lisosoma e utilizzate dalla cellula. Digestione intracellulare di materiale o organuli cellulari invecchiati in questo caso il lisosoma ingloba una struttura cellulare per esempio un mitocondrio, forma così un autofagosoma in cui l'azione dei suoi enzimi demolisce la struttura inglobata. I prodotti della digestione quindi riassorbite e utilizzati dalla cellula. Autolisi della cellula. La membrana del lisosoma in questo caso si rompe per sopraggiunte situazioni patologiche o a causa fisiologiche (azione ormonale) gli enzimi quindi risultano liberi di agire sulle strutture cellulari e quindi le digeriscono, per questa loro azione i lisosomi sono chiamati anche sacchetti suicidi.
Il nucleo è il centro di controllo genetico della cellula
Questo è l'organulo tipico della cellula eucariote, delimitato da una doppia membrana e presenta al suo interno proteine e acidi nucleici (soprattutto DNA, che rappresenta il materiale genetico, e RNA). Il nucleo può essere osservato sotto forma di cromatina, sostanza visibile attraverso la preparazione con coloranti per microscopia luce o attraverso la microscopia elettronica, che permette di apprezzarne la struttura e l'organizzazione. É l'organulo con dimensioni maggiori rispetto agli altri organuli. Il nucleo costituisce il centro di controllo funzionale dell'informazione genetica della cellula. Al suo interno si svolgono:
la replicazione del DNA (autoduplicazione) prima della divisione cellulare,
la trascrizione (produzione) dei diversi tipi di RNA che verranno utilizzati nella sintesi proteica (traduzione dell'informazione genetica)
Struttura e organizzazione: tutte le cellule eucariotiche presentano un solo nucleo, tranne alcuni tipi di cellule quali i globuli rossi del sangue che non ce l'hanno, a altri tipi che ne possono avere due (come le cellule del fegato). Ogni nucleo si origina per divisione di un altro nucleo preesistente, quando sta per avvenire la divisione cellulare. A questa regola c'è un'importante eccezione: dopo la fecondazione il gamete maschile e quello femminile si fondono e così anche i loro nuclei per dare origine il primo nucleo del futuro individuo.
Durante il ciclo cellulare il nucleo cambia aspetto. Per descriverne la struttura si tiene conto di una particolare fase della vita cellulare, chiamata interfase cioè periodo che separa una fase di attività intensa del metabolismo cellulare (sintesi proteica, duplicazione del DNA) alla divisione cellulare comprendente la mitosi. Durante l'interfase il nucleo è visibile e si presenta sotto forma sferica le cui dimensioni sono in stretta relazione con le dimensioni della cellula. Mentre durante le altre fasi del ciclo cellulare il nucleo non è più visibile tende a sparire per poi ricomporsi dopo avvenuta divisione cellulare. È proprio durante la interfase che si possono riconoscere le sue parti costituenti (membrana, nucleoplasma e nucleolo)
Le funzioni della membrana nucleare:
separare le attività del genoma (patrimonio genetico) quali la replicazione (mantenimento dell'informazione genetica tra una generazione e quella successiva), la trascrizione del DNA (passaggio dell'informazione genetica tra il nucleo e il citoplasma) dalle funzioni che avvengono nel citoplasma tra cui la traduzione dell'informazione genetica nella produzione di proteine
regolare la comunicazione, cioè il trasporto di molecole, tra il nucleo e il citoplasma
Struttura della membrana nucleare: due membrane concentriche e parallele (interna ed esterna) separate da un sottile spazio perinucleare. Ogni membrana presenta la stessa organizzazione trilaminare delle altre membrane cellulare (doppio strato lipidicoproteico) ed è dotata di numerose aperture (3.000-4.000) dette pori nucleari. La membrana nucleare è molto selettiva: evita il passaggio casuale di materiali tra citoplasma e nucleo, regolando con molta efficienza ciò che può entrare al di dentro. Passano attraverso i pori nucleari, in ingresso:
i nucleotidi necessari alla sintesi degli acidi nucleici
le piccole molecole proteiche regolatrici dell'attività genica.
in uscita:
gli RNA e i ribosomi sintetizzati dal nucleo
La membrana esterna è in continuità con il reticolo endoplasmatico, mentre la membrana interna poggia su una rete di particolari proteine, che svolge funzione di sostegno. A livello di ciascun poro le due membrane si fondono. Il poro è in definitiva costituito da un canale cilindrico che mette in comunicazione il citoplasma della cellula con lo spazio interno al nucleo. Il trasporto da e per il nucleo è di due tipi:
passivo permette il passaggio libero, senza dispendio di energia, dei nucleosidi necessari alla sintesi di DNA e RNA, le molecole proteiche più piccole coinvolte nella condensazione e nella trascrizione del DNA e gli ioni che aiutano ad equilibrare la concentrazione di essi fra l'interno del nucleo e il citoplasma
attivo per le molecole proteiche più grandi o le molecole di RNAm
All'interno del nucleoplasma vi è una matrice gelatinosa in cui è sospesa la cromatina, cioè il DNA e le proteine associate. La matrice nucleare si presenta come una ordinata architettura in cui è permesso il regolare ed efficiente svolgimento della replicazione e della trascrizione del DNA, in quanto le fibre di cromatina non sono dispose casualmente, ma ognuna occupa un preciso dominio spaziale. Quando sta per avvenire la divisione cellulare la cromatina si addensa e si organizza in strutture individuali, a forma di piccoli bastoncini, detti cromosomi. Ogni specie vivente è caratterizzata dall'avere numero e forma di questi bastoncini specifici determinati e fissi.
Un cromosoma è una molecola di DNA molto grande che contiene una serie di geni, complessata con diverse proteine. Come si vede dalla figura ogni cromosoma è costituito da due subunità detti cromatidi, cioè due filamenti identici di DNA uniti in un punto detto centromero. Nella parte centrale del nucleo c'è una struttura molto densa e sferica formata da due zone:
una fibrillare più interna che contiene il DNA
l'altra granulare contiene RNA (RNA ribosomiale o r-RNA) e proteine importate dal citoplasma
Il nucleolo è responsabile della sintesi dell'RNA ribosomiale ed è ricco di ribosomi in varie fasi della oro formazione. I ribosomi lasciano il nucleo per dirigersi verso il citoplasma, dove avviene l'assemblaggio nelle due subunità mature per partecipare alla sintesi delle proteine.
Il mitocondrio preleva l'energia chimica presente negli alimenti
camera esterna matrice mitocondriale
membrana esterna liscia
membrana interna più estesa si
ripiega più volte su se stessa fino a
formare le creste mitocondriali
Organulo generalmente di forma allungata o granulare ubiquitario in tutte le cellule eucariotiche (descritto per la prima volta nel 1894). L'intero corredo di mitocondri della cellula si chiama condrioma. Anche per questo organulo la microscopia elettronica ne ha permesso l'osservazione dell'ultrastruttura. Per comprendere la complessità dell'insieme delle funzioni metaboliche del mitocondrio è stato inoltre necessario lo studio combinato con la biochimica molecolare. Diamo alcune informazioni sulla struttura: organulo delimitato da due membrane: una esterna e l'altra interna separate da uno spazio detto camera esterna. La membrana interna delimita lo spazio centrale detto camera interna, occupata dalla cosiddetta matrice mitocondriale. Nella matrice ci sono ribosomi, una o più molecole di DNA circolare (DNA mitocondriale), sono stati identificati, attraverso la biochimica, anche t-RNA (di trasporto) e m-RNA (messaggero). La membrana interna è molto più estesa di quella esterna e forma diverse pieghe ed estroflessioni chiamate creste mitocondriali che si affacciano nella camera interna. Il DNA e l'RNA del mitocondrio formano un sistema genetico autonomo che, con la presenza di ribosomi, conferisce all'organulo una capacità autonoma di sintetizzare alcune proteine di cui ha bisogno per svolgere le diverse funzioni di cui è deputato. Per il completamento delle funzioni necessita di altre proteine, la maggior parte, che giungono dal citoplasma.
Il mitocondrio è in grado di svolgere molteplici funzioni:
la più importante tra esse è legata al processo di respirazione aerobica, cioè all'insieme di reazioni con le quali la cellula effettua l'ossidazione completa dei materiali organici nutritivi, ciò permette di estrarre energia dai substrati organici che gli arrivano, producendo un gradiente ionico che viene successivamente utilizzato per sintetizzare adenosintrifosfato (ATP nucleotide con tre gruppi fosfati) detta moneta energetica biologica.
Qui si da qualche cenno all'importante serie di reazioni ossidative della respirazione cellulare che riprenderemo in modo più dettagliato successivamente. Nella matrice mitocondriale ci sono gli enzimi e i coenzimi necessari nelle diverse tappe del ciclo di Krebs, con cui le ossidazioni iniziate nel citoplasma (glicolisi) si completano con produzione di:
CO2 che viene liberata
ioni H+ ed elettroni e- vengono assunti da una serie di complessi enzimatici (catena respiratoria) situati nelle creste e trasferiti, attraverso una ordinata sequenza di ossidoriduzioni fino all'O2 per formare acqua (H2O)
Durante le ossidoriduzioni si libera energia che viene accumulata in molecole di ATP in un processo chiamato fosforilazione ossidativa. I complessi enzimatici della catena respiratoria sulle creste mitocondriali in funzione delle reazioni che catalizzano.
I mitocondri hanno un'enorme importanza per le cellule quali centrali energetiche. Una cellula demolendo una molecola di glucosio, ricava solo due molecole di ATP con la glicolisi, che avviene nel citoplasma, ma ne estrae in totale 36 se la demolizione prosegue a livello dei mitocondri con il processo di respirazione aerobica.
Nella fase di divisione cellulare i mitocondri si ripartiscono tra le due cellule figlie e durante l'interfase il loro numero aumenta per divisione dei mitocondri preesistenti.
Particolari strutture delle cellule vegetali Vacuoli mantenimento omeostasi
Questi particolari organuli delle cellule vegetali sono stati osservati molto precocemente a causa delle loro enormi dimensioni rispetto al volume totale della cellula di appartenenza. Nelle prime osservazioni essi furono ritenuti cavità vuote, difatti vennero chiamati vacuoli dal latino vacuus = vuoto, ma a dispetto delle apparenze essi non sono vuoti e si comportano come organuli.
Sono organuli solitamente di forma globosa e si presentano come una grossa vescicola delimitata da una singola membrana semipermeabile di natura lipoproteica, detta tonoplasto.
Il tonoplasto è costituito da una sola membrana frequentemente asimmetrica, perché la componente proteica volta verso l'interno della cisterna risulta maggiormente sviluppata rispetto a quella che rivolge la faccia verso il citoplasma. Nelle cellule vegetali adulte (d) di norma si rinviene un unico grande vacuolo centrale, che può occupare anche più del 90% del volume cellulare.
Nelle cellule meristematiche (a), si trovano piccoli e numerosi vacuoli che, poi, nelle cellule differenziate adulte confluiranno nel vacuolo unico. Con l'avvio della distensione cellulare e con il differenziamento il numero dei vacuoli aumenta, così come le loro dimensioni (b e c), a causa dell'assorbimento dell'acqua da parte della cellula. Molti vacuoli si saldano tra loro e alla fine della distensione, quando la cellula ha raggiunto le sue dimensioni caratteristiche, nella maggior parte dei casi ha un solo grande vacuolo centrale (d) attorno al quale il citoplasma rimane a costituire un piccolo velo. Con l'aumento delle dimensioni del vacuolo, la cellula vegetale può attuare una notevole distensione, raggiungendo dimensioni ragguardevoli, superiori a quelle della maggior parte delle cellule animali. La funzionalità cellulare risulta favorita dallo spostamento del citoplasma alla periferia, perché con questo aggiustamento rimangono proporzionati i rapporti tra volume del plasma e superficie di scambio.
Le funzioni:
evita la formazione di spazi vuoti
spinge il citoplasma verso l'esterno della cellula facilitando gli scambi metabolici
rappresenta un sistema di escrezione dei rifiuti
regola l'omeostasi, funzionando come osmometro, mediante variazioni di concentrazione del succo vacuolare
organulo di riserva (acqua e varie sostanze)
funzione vessillare in quanto concorre alla colorazione di fiori, frutti e altre parti vegetali per la presenza di pimenti
I vacuoli prendono origine dal GERL (insieme di parti comuni al reticolo endoplasmatico e apparato del Golgi) e sono ripieni di un succo vacuolare acquoso, di composizione chimica molto varia sia nei diversi gruppi vegetali sia, nello stesso individuo, secondo la specializzazione funzionale delle sue cellule. Il succo vacuolare può essere costituito da soluzioni vere, soluzioni colloidali e concrezioni solide di vario genere. Nel vacuolo vengono immagazzinate sostanze di riserva, prodotti secondari e scorie del metabolismo. La composizione chimica del succo vacuolare, il cui pH è sempre acido (4-5):
velo. Con l'aumento delle dimensioni del vacuolo, la cellula vegetale può attuare una notevole distensione, raggiungendo dimensioni ragguardevoli, superiori a quelle della maggior parte delle cellule animali. La funzionalità cellulare risulta favorita dallo spostamento del citoplasma alla periferia, perché con questo aggiustamento rimangono proporzionati i rapporti tra volume del plasma e superficie di scambio.
Le funzioni:
evita la formazione di spazi vuoti
spinge il citoplasma verso l'esterno della cellula facilitando gli scambi metabolici
rappresenta un sistema di escrezione dei rifiuti
regola l'omeostasi, funzionando come osmometro, mediante variazioni di concentrazione del succo vacuolare
organulo di riserva (acqua e varie sostanze)
funzione vessillare in quanto concorre alla colorazione di fiori, frutti e altre parti vegetali per la presenza di pimenti
I vacuoli prendono origine dal GERL (insieme di parti comuni al reticolo endoplasmatico e apparato del Golgi) e sono ripieni di un succo vacuolare acquoso, di composizione chimica molto varia sia nei diversi gruppi vegetali sia, nello stesso individuo, secondo la specializzazione funzionale delle sue cellule. Il succo vacuolare può essere costituito da soluzioni vere, soluzioni colloidali e concrezioni solide di vario genere. Nel vacuolo vengono immagazzinate sostanze di riserva, prodotti secondari e scorie del metabolismo. La composizione chimica del succo vacuolare, il cui pH è sempre acido (4-5):
Acqua
Glucidi: monosaccaridi (glucosio, fruttosio), disaccaridi (saccarosio, maltosio), polisaccaridi (inulina)
Sempre presenti soluzioni saline diluite di sali inorganici (tipo e concentrazione dipendono dall'ambiente) in cui i cationi più abbondanti sono Ca2+, K+, Mg2+ e Na+, mentre gli anioni più rappresentati sono ione solfato, fosfato, nitrato e cloro, ecc.)
Sali di acidi organici: acido citrico (nei frutti immaturi), acido malico, acido succinico, acido ossalico, ecc., spesso in cristalli
Amminoacidi: arginina, lisina
Flavonoidi:pigmenti rossi e blu (antociani) o gialli (flavoni)
Alcaloidi:sostanze basiche probabilmente con funzione di difesa (morfina, cocaina, nicotina, stricnina)
Altri metaboliti: tannini, glucosidi, resine, gomme, oli essenziali, ecc.
Plastidi processi di fotosintesi e di accumulo sostanze di riserva
Questi organuli sono propri delle cellule vegetali autotrofe, considerati un'entità biologica unica, perché a partire da un protoplastidio comune (forma giovanile del plastidio, presente nei tessuti meristematici primari) si possono evolvere in maniera diversa (organuli definitivi morfologicamente e fisiologicamente differenti: cloroplasti, cromoplasti e leucoplasti), a seconda delle condizioni ambientali e dei fattori interni di determinazione a cui sono sottoposti durante le varie fasi di sviluppo vegetale.
La struttura dei plastidi è semplice in quanto sono delimitati da due membrane (interna ed esterna) che racchiudono uno stroma uniforme, privo di qualsiasi struttura lamellare. Forma ellissoide e dimensioni pari a circa 1 μm quelli non differenziati e pari a 5-10 μm quelli differenziati. Nello stroma (fase amorfa) si trovano il DNA, l'RNA (organizzato anche in ribosomi), e una serie di enzimi che li rende capaci di sintetizzare proteine, lipidi e anche amido, se ricevono gli zuccheri necessari dal resto della cellula. Quindi come per il mitocondrio ci troviamo di fronte ad un organulo semi autonomo. Nelle cellule vegetali delle piante superiori si osservano tre tipi di plastidi: cloroplasti, cromoplasti e leucoplasti. Questi tre tipi, come abbiamo già detto, derivano dai protoplastidi non differenziati presenti nei tessuti meristematici primari. Possono altresì formarsi per divisione dei plastidi differenziati esistenti, oppure per trasformazione di un tipo in un altro. Sono quasi sempre in attiva duplicazione.
I plastidi si differenziano in cloroplasti solo in presenza di luce, questo fattore è indispensabile, ma non è l'unico perché entrano in gioco altri fattori di determinazione che regolano o bloccano tale processo. Non sempre la luce determina la differenziazione dei plastidi in cloroplasti, infatti le cellule dell'epidermide non presentano cloroplasti, anche se tale tessuto si sviluppa in piena luce, perché in questo caso prevale la determinazione fisiologica che prevede per le cellule una funzione protettiva e non fotosintetizzante; mentre le cellule guardia degli stomi sono ricchi di cloroplasti che permettono la fotosintesi, processo importante per la regolazione dell'apertura degli tomi e per il conseguente scambio gassoso con l'atmosfera. I protoplastidi delle cellule del meristema primario dell'apparato radicale normalmente non si differenziano in strutture fotosintetizzanti, ma si trasformano in leucoplasti o cromoplasti.
Leucoplasti così chiamati per l'assenza di pigmenti e perciò incolori. Sono organuli che accumulano sostanze di riserva e, a seconda della natura chimica di queste sostanze, sono distinti in:
amiloplasti si trovano nei parenchimi amiliferi di riserva, cioè accumulano amido secondario: frutti, semi, fusti, radici. La loro caratteristica biologica più evidente è la capacità di sintesi di amido e di idrolisi di questo in glucidi semplici. Gli amiloplasti sono diffusi in tessuti adulti che non vengono colpiti direttamente dalla luce, presentano una struttura relativamente semplice, circondati da una doppia membrana, la più interna differenzia pochi ed incompleti strati lamellari. Gli amiloplasti contengono ingenti quantità di enzimi, quelli che presiedono alla sintesi degli zuccheri e alla loro polimerizzazione. Vi è come in tutti i plastidi anche il DNA e l'RNA, lo stroma è occupato in gran parte dalle sostanze di riserva (amido secondario) sotto forma di grossi granuli molto stabili e di dimensioni variabili a volte così grandi da occupare quasi tutto lo stroma dell'organulo. La forma dei granuli è un carattere diagnostico per il riconoscimento della specie. Il centro di formazione del granulo si chiama ilo e questo può essere unico (granuli semplici es. patata , grano) o più di uno nello stesso amiloplasto e si costituiranno granuli complessi (riso, avena..)
elaioplasti che accumulano plastoglobuli lipidici, tipici di certe famiglie (es. Cactacee. Liliacee, Cucurbitacee)
proteoplasti accumulatori di proteine
Cromoplasti sono plastidi adulti ricchi di pigmenti, perciò colorati, ma non fotosintetici. Fra i pigmenti sono prevalenti i carotenoidi (fino al 30% del peso secco dell'organulo) e assumono vari colori nella gamma dei gialli e dei rossi. Sono molto presenti in fiori, frutti e foglie e insieme agli antociani che sono disciolti nei vacuoli conferiscono vivaci colorazioni agli organi vegetali (funzione vessillare per attirare insetti pronubi e gli animali frugivori). Questi organuli sono privi di clorofilla, quindi fotosinteticamente inattivi. Anche essi sono delimitati da una doppia membrana che racchiude lo stroma in cui sono presenti vescicole, lamelle, cristalli, tubuli... Sono presenti lipidi (58%), proteine (22%), RNA (3%) e forse anche DNA e RNA ribosomiale. Derivano dai protoplastidi oppure dai cloroplasti per eliminazione della clorofilla. La derivazione dai cloroplasti è facilmente riconoscibile durante la maturazione dei frutti o durante la senescenza delle foglie e si accompagna a un riassorbimento della clorofilla e a un riassetto del sistema di membrane stromatiche e granali.
I cloroplasti trasformano l'energia solare in energia chimica
I cloroplasti svolgono il processo della fotosintesi e nelle piante superiori rappresentano uno degli organuli di maggiore dimensione con lunghezza di 5-10 μm e diametro di 1-3 μm. Il fattore luce condiziona enormemente la possibilità dei protoplastidi di differenziarsi in cloroplasti. Questi plastidi sono particolarmente abbondanti nei parenchimi clorofilliani delle foglie e dei giovani fusti di consistenza erbacea. Sono capaci di dividersi autonomamente, indipendentemente dalla mitosi cellulare. Un cloroplasto maturo ha la seguente composizione: 40-60% del p.s. proteine; 20-30% lipidi, il 5-10% da clorofille (75% clorofilla a e 25% da clorofilla b), 2-4% da carotenoidi, RNA circa 6%, DNA 0,5%. La parte restante (7-10%) è rappresentato dalla parte inorganica di sali minerali (K, Ca, Mg, Na e Mn).
Struttura nello stroma dell'organulo avviene l'elaborazione dei pigmenti, in particolare di clorofille e carotenoidi, contemporanea alla formazione delle lamelle. Grazie ad un proprio DNA lo stroma è in grado di sintetizzare proteine, in particolare quelle strutturali dell'apparato fotosintetico. La membrana interna del plastidio si invagina in numerosi punti formando dei sacculi appiattiti che si staccano dalla membrana stessa e si fondono fra loro dando origine a formazioni lamellari lunghe quanto l'organulo, dette tilacoidi stromatici. In seguito in alcuni punti si registra l'impilamento di un certo numero di vescicole appiattite che costituiscono nel loro insieme un granum, con diametro medio di 0,3-2 μm. I grana costituiscono un mezzo per aumentare la superficie interna della struttura fotosintetizzante e rappresentano le strutture dei cloroplasti adibiti alla cattura dell'energia solare.
Parete cellulare
La cellula vegetale è delimitata, oltre che dalla membrana plasmatica, anche da uno strato extraprotoplasmatico, molto organizzato, di natura polisaccaridica, chiamato parete. Questo rivestimento esterno è tipico dei vegetali, ma è possibile trovarlo, con parziale variazione nella composizione chimica, anche nei batteri e nei funghi. La parete cellulare dei vegetali si presenta come una scatola rigida che circonda la cellula (vedi figure nella pagina precedente). Le funzioni della parete:
conferimento della rigidità a certe strutture
mantenimento della forza meccanica e aumento della solidità di certi organi
controllo della distensione cellulare
controllo del trasporto intercellulare
protezione da microrganismi patogeni
produzione di molecole segnale, specialmente in relazione ad attacchi di patogeni
accumulo di sostanze organiche
regolazione flusso d'acqua in ingresso e in uscita
La parete cresce con l'aumentare del volume cellulare, contiene enzimi e, a volte, sono presenti dei ribosomi, per tali ragioni è considerato un organulo cellulare che interagisce con il resto della cellula.
L'accrescimento della parete primordiale originata dalla divisione della cellula madre comporta la costituzione della parete primaria (si origina dagli strati esterni, in cui sono presenti fibrille cellulosiche, mentre la zona centrale, pectica, si trasforma in lamella primaria, con funzione cementante (salda assieme cellule adiacenti). I vegetali depongono la loro parete a strati. Ciò permette di riconoscere tre zone:
lamella mediana zona centrale costituita essenzialmente da pectine
parete primaria formata da una matrice in cui è disperso materiale fibrillare. La matrice è costituita in percentuale elevata da acqua (60%), cellulosa, emicellulose, proteine e lipidi
Se la cellula non è meristematica (soggetta a divisioni), ma si specializza in funzioni particolari, alla parete primaria si appone, dall'interno, la parete secondaria, caratterizzata da un forte incremento percentuale della componente cellulosica, mentre diminuisce la proporzione delle pectine, delle emicellulose, delle proteine. Anche nella parete secondaria si riconosce una matrice, la cui quantità è però molto scarsa, e un sistema fibrillare cellulosico che prevale su tutto il resto.
La cellulosa è la molecola organica più abbondante sulla Terra. Essa si trova in varie forme (carta, legno, materie plastiche cellulosiche, fibre tessili), si trova alla base di numerose attività economiche umane. È un polimero del glucosio.
La parete secondaria è normalmente stratificata e si accresce per la deposizione centripeta di lamelle concentriche. è tipica delle cellule che hanno terminato il loro processo di distensione e differenziamento. Le variazioni di composizione rispetto alla parete primaria dipendono dalla comparsa di nuove sostanze (lignina, cere e cutine, suberina) e dalla variazione percentuale di quelle precedentemente presenti. In certi elementi del legno (come fibre e tracheidi delle conifere) si possono riconoscere tre strati, ciascuno costituito di un numero elevato di lamelle, in cui microfibrille sono alternativamente orientate in senso opposto. La struttura stratificata e il, diverso orientamento delle microfibrille, che risultano disposte a formare un traliccio, spiegano la particolare resistenza delle pareti secondarie.
La particolare organizzazione strutturale della parete secondaria, lasciando liberi interstizi, fa in modo che all'interno possano depositarsi componenti molecolari diverse da quelle pectocellulosiche ed emicellulosiche. Si verificano così dei processi di modificazione della parete, connessi con la differenziazione cellulare. A seconda delle sostanze che vengono deposte, le modificazioni prendono i seguenti nomi: cutinizzazione, suberificazione, lignificazione, mineralizzazione, pigmentazione....
Organuli eucarioti e loro funzioni
Funzione generale: assemblaggio
Nucleo Sintesi di DNA; sintesi di RNA; assemblaggio di subunità ribosomiali (nel nucleolo)
Ribosomi sintesi di poplipeptidi (proteine)
Reticolo endoplasmatico rugoso sintesi delle proteine di membrana, delle proteine da secernere e degli enzimi idrolitici; formazione delle vescicole di trasporto
Reticolo endoplasmatico liscio sintesi di lipidi; metabolismo dei carboidrati nelle cellule del fegato; demolizione di sostanze nocive nelle cellule del fegato; immagazzinamento di ioni calcio
Apparato del Golgi rielaborazione, deposito temporaneo e trasporto di macromolecole; formazione di lisosomi e di vescicole di trasporto
Funzione generale: demolizione
Lisosomi digestione delle sostanze nutritive, dei batteri e degli organuli danneggiati; distruzione di certe cellule durante lo sviluppo embrionale
Vacuoli digestione (come i lisosomi); immagazzinamento di sostanze chimiche; aumento delle dimensioni cellulari; bilancio idrico
Funzione generale: trasformazioni energetiche
Cloroplasti conversione di energia luminosa in energia chimica contenuta negli zuccheri
Mitocondri conversione di energia chimica degli alimenti racchiusa in molecole di ATP
Funzione generale: sostegno, movimento
Citoscheletro mantenimento della forma cellulare; ancoraggio per gli organuli; movimento degli organuli nelle cellule; movimento cellulare
Parete cellulare mantenimento della forma cellulare e sostegno scheletrico; protezione delle superfici; fissaggio delle cellule nei tessuti
Fonte: http://www.giottoulivi.it/studioinrete/scienze_Santoro/strutture&funzioni_cellule.doc
Prof Michele Santoro
Cellule animali e cellule vegetali
Il ciclo cellulare
Il ciclo cellulare si divide in tre fasi:
- L’interfase;
- La mitosi;
- La citodieresi;
L’INTERFASE
L’interfase è la fase preparativa alla divisione cellulare; essa può essere divisa principalmente in tre sottofasi: le fasi G1, S, G2.
- Fase G1: la cellula raddoppia le sue dimensioni. Aumenta anche il numero d’enzimi, ribosomi, centrioli, mitocondri e delle altre strutture endoplasmatiche. La cellula inoltre crea nuove strutture come microtuboli e filamenti d’actina. L’apparato di Golgi e altre strutture membranose sono create dal reticolo endoplasmatico. Un fatto interessante è che i mitocondri e i cloroplasti vengono prodotti solo da loro simili. Infatti essi contengono un cromosoma indipendente da quelli della cellula.
- Fase S: questa fase, la più importante dell’interfase ha la funzione di sintetizzare molte proteine associate al DNA.
- Fase G2: è la fase conclusiva dell’interfase; ha il compito di spiralizzare i cromosomi presenti nel nucleo. In questa fase inoltre i due centrioli si dispongono alle due estremità opposte della parete nucleare. Infine vengono riunite tutte le strutture necessarie per donare alle cellule figlie un corredo cromosomico completo.
La mitosi
La mitosi si divide principalmente in 4 parti: profase, metafase, anafase, telofase.
- Profase: i cromosomi si spiralizzano. Ogni cromosoma è formato da 2 cromatidi uniti insieme nella regione mediana detta centromero; al centromero sono anche attaccate 2 strutture proteiche chiamate cinetocori. Durante la mitosi inoltre i centrioli si separano e si viene lentamente a formare un fuso di microtuboli. Le fibre che compongono questo fuso possono essere divise principalmente in tre tipi: fibre polari, che hanno il compito di unire gli estremi del fuso con la parte centrale, le fibre del cinetocore che hanno il compito di congiungere gli estremi del fuso con i cinetocori dei cromosomi, e, nelle cellule in cui sono presenti centrioli, l’aster, che lega gli estremi del fuso (poli) con la membrana cellulare.
- Metafse: grazie a movimenti delle fibre del cinetocore, i cromosomi vengono allineati nella regione mediana del fuso.
- Anafase: i due cromatidi si separano. Ciascun cromatida trascinato dalle fibre del cinetocore, viene trasportato in un estremo della cellula.
- Telofase: ultima parte della mitosi: durante questa fase il fuso si disperde, si riforma la membrana nucleare, i cromosomi riprendono la loro forma originaria e vicino alla coppia di centrioli rimasta se en inizia a formare un'altra.
La citodieresi
La citodieresi è il processo tramite il quale si divide il citoplasma. Questa ha inizio durante la telofase e segue 2 procedimenti differenti a seconda che la cellula sia animale o vegetale.
- Citodieresi nelle cellule animali: durante la telofase la membrana cellulare inizia a restringersi in prossimità della zona equatoriale del fuso finché non si ha una strozzatura divide definitivamente la cellula madre in 2 cellule figlie.
- Citodieresi nelle cellule vegetali: in queste cellule l’apparato di Golgi produce una serie di vescicole contenenti polisaccaridi. Queste si vanno a depositare sulla linea mediana, si fondono insieme e formano una parete chiamata piastra cellulare. Quando questa lamella è completata ogni cellula costruisce la sua membrana cellulare terminando cosi il processo di divisione.
Scritto da: Michele Monti
http://riappunti.net/biologia/Il_ciclo_cellulare.doc
Cellule animali e cellule vegetali
Visita la nostra pagina principale
Cellule animali e cellule vegetali
Termini d' uso e privacy